



 Éd. 1971-1976
Éd. 1971-1976testicule
Glande génitale mâle, lieu de fabrication des cellules sexuelles mâles, ou spermatozoïdes.
L’origine du testicule
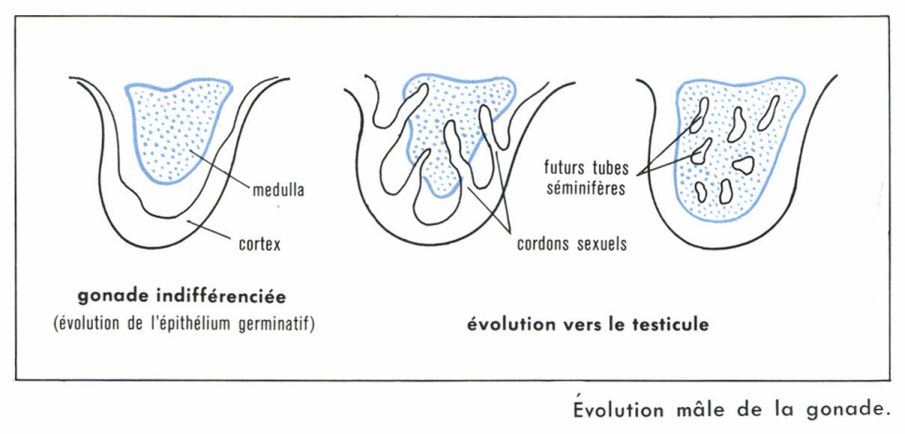
Une partie de l’épithélium péritonéal est colonisée par les cellules de la lignée germinale. Elle deviendra l’épithélium germinatif, d’où seront issus cortex et medulla de la gonade indifférenciée. Cette évolution, qui se produit aussi bien chez le garçon que chez la fille, se déroule pendant les cinquante premiers jours de la vie fœtale. Au-delà de ce temps, une unique poussée de cordons sexuels à partir du cortex au sein de la medulla va conduire progressivement à la formation des tubes séminifères. Entre ces tubes, la persistance d’un abondant tissu interstitiel va être à l’origine de la sécrétion d’un hormone mâle, la testostérone. Le testicule est alors constitué ; d’intra-abdominal, il deviendra extra-abdominal en descendant dans les bourses. Cependant, la spermatogenèse ne débutera qu’après la puberté.
La structure du testicule
Chez les Vertébrés supérieurs, dont l’Homme, les deux testicules se présentent comme un ensemble de tubes séminifères (tubes en cul-de-sac) collectés par d’autres canaux (tubuli recti) qui s’anastomosent pour former un réseau de tubules (rete testis). Ces derniers quittent le testicule (vasa efferentia) et se jettent dans un tube unique, mais très contourné, l’épididyme. Entre les tubes séminifères, on trouve un tissu conjonctif à grandes cellules (cellules de Leydig). Ce tissu interstitiel est à l’origine de la sécrétion d’hormones mâles.
À la suite de l’épididyme débute le canal évacuateur des spermatozoïdes (canal déférent, ou spermiducte), dont la fonction essentielle est de conduire les cellules sexuelles mâles jusqu’à l’urètre, voie commune d’évacuation de l’urine et du sperme. Le schéma varie quelque peu chez les autres Vertébrés (cystes ou ampoules des Amphibiens), mais surtout au niveau des relations glande sexuelle-appareil urinaire, ce qui s’explique par l’évolution commune des deux appareils.
La variabilité, plus grande encore chez les Invertébrés (ampoules séminales des Insectes par exemple), cache une unité profonde de fonction : la genèse des spermatozoïdes, ou spermatogenèse.


Les mécanismes de la spermatogenèse
La découverte (chez l’Ascaris) de la spermatogenèse ne remonte qu’à 1880 (époque des recherches du biologiste allemand Oskar Hertwig [1849-1922]). Elle se décompose en deux périodes : la réduction chromatique, ou méiose, et la différenciation cellulaire, qui lui fait suite et qui conduit à la formation de cette cellule très particulière qu’est le spermatozoïde.
La méiose s’inscrit dans le cadre plus général du cycle chromosomique, auquel est soumis tout individu. Comme dans le cas de l’ovogenèse, chez l’animal mâle, c’est au moment de la spermatogenèse qu’on constate que certaines cellules, dites « de la lignée germinale », voient leur nombre de chromosomes passer de 2n à n. Ces cellules à 2n chromosomes, ou spermatogonies, vont s’accroître légèrement et donner naissance aux spermatocytes de premier ordre (spermatocytes I), qui vont subir la mitose réductionnelle de la méiose, donnant chacun naissance à deux spermatocytes II (ou de second ordre) à n chromosomes. Ces spermatocytes II subiront à leur tour la mitose équationnelle, qui les divisera en deux spermatides à n chromosomes.
C’est à ce moment que se situe l’originalité de la spermatogenèse, car, d’une cellule, haploïde certes, mais banale dans sa structure, on va passer à une cellule très différenciée, le spermatozoïde.
Les chromosomes nucléaires se présentent sous forme de fins filaments homogènes, dont l’A. D. N. résiste à l’action de l’enzyme désoxyribonucléique (l’A. D. N. représente d’ailleurs 20 p. 100 du poids sec total du spermatozoïde).
C’est au niveau du cytoplasme que les transformations sont les plus profondes. L’appareil de Golgi élabore des granules qui donneront naissance à une structure qui coiffe le noyau, l’acrosome.
Les deux centrioles du centrosome viennent se placer à l’opposé de l’acrosome par rapport au noyau. L’un d’eux, le plus éloigné du noyau, élabore un flagelle, autour duquel s’allonge le reste du cytoplasme. À proximité du noyau, dans ce reste de cytoplasme, se forme une gaine de mitochondries autour du flagelle.
Acrosome, permettant la pénétration du spermatozoïde dans le gamète femelle, centrioles, dirigeant l’établissement de la dicentrie, et flagelle, assurant la mobilité du spermatozoïde, sont autant d’organites qui contribuent largement à la fécondation.
Spermatogenèse, sperme et fécondité
Chez les Mammifères, les spermatozoïdes mesurent entre 40 et 250 μ (Homme : 53 μ). Leur longueur peut atteindre quelques millimètres chez certaines espèces (Pontocypris monstrosa, Crustacé ostracode : 6 mm). De toute façon, leur taille est généralement beaucoup plus faible que celle de l’ovule (le spermatozoïde de Taureau a un volume égal au 1/20 000 du volume du gamète de la Vache).
La production de spermatozoïdes par le testicule est variable. Elle est de 3 . 106 gamètes chez le Taureau et de 14 . 106 chez le Lapin par gramme de testicule et par jour.
L’hormone folliculostimulante de l’hypophyse (F. S. H.) active le fonctionnement des tubes séminifères et, de ce fait, la spermatogenèse.
En raison des relations hypothalamo-hypophysaires, on conçoit que le système nerveux puisse contrôler la genèse des cellules mâles. Les phénomènes extérieurs, tels que les afférences lumineuses (longueur du jour), peuvent avoir une influence suffisante pour expliquer l’activité périodique de la spermatogenèse chez de nombreuses espèces animales.
À la sortie du testicule, la liqueur spermatique est constituée par les seuls spermatozoïdes. Par contre, au moment de l’éjaculation, le sperme contient, outre les cellules sexuelles, les sécrétions des glandes annexes de l’appareil génital mâle : vésicules séminales et prostate. Le facteur de dilution est d’au moins trois fois. Conséquence : le nombre de spermatozoïdes par millimètre cube de sperme varie non seulement en fonction de la production testiculaire, mais aussi des sécrétions glandulaires. Cette variabilité se retrouve à travers les espèces animales :
On a constaté que les spermatozoïdes issus directement du testicule n’étaient pas fécondants, leur motilité étant pratiquement nulle. Ils n’acquièrent leur complète activité qu’après passage dans l’épididyme et grâce aux sécrétions des vésicules séminales et de la prostate. Le parcours est d’environ 6 m chez l’Homme (40 m chez le Taureau).