sensation (suite)
• Caractères spatiaux et temporels. Certaines catégories de sensations, principalement tactiles, visuelles et ultrasonores ou électro-esthésiques (v. écholocation et électricité animale), sont affectées d’un « signe local » extrêmement précis. À partir de ces signes locaux peut alors s’édifier une structure complexe, le « champ sensoriel », représentatif de certaines propriétés de l’espace qui entoure l’animal ; champ visuel, champ ultrasonore, etc. La finesse d’analyse de ce champ dépend d’un seuil de discrimination spatiale (pour l’œil humain, angle d’environ 1′). Il est évident que les capacités de survie d’un animal chasseur (ou chassé) dépendent directement de ses performances dans ce domaine : l’Oiseau insectivore qui donne des coups de bec toujours à côté de sa proie a toutes chances de mourir de faim !
Quant au « signe temporel », c’est la reconnaissance des rythmes d’apparition et de disparition des stimuli, de leur accroissement ou de leur diminution plus ou moins rapide, bref, du profil temporel de la courbe représentative du phénomène. Ce signe temporel joue un rôle de tout premier plan dans la reconnaissance mutuelle des individus au sein d’une même espèce (couples d’Oiseaux, familles de Pingouins) et dans l’édification du champ ultrasonore des Chauves-Souris, ou encore dans la capture des proies chez les Grenouilles, qui ne réagissent qu’au passage rapide d’une ombre. Le vaste domaine de la reconnaissance des mouvements des proies repose entièrement sur l’existence de ces sensibilités aux changements rapides. Bien entendu, on bute, ici encore, sur des seuils : une discontinuité visuelle de trop haute fréquence n’est plus perçue, on a atteint la « fréquence critique fusion » ; on sait que c’est là le principe du cinéma.
Analyse des mécanismes
La fonction des systèmes sensoriels
La fonction des systèmes sensoriels est de fournir au système nerveux central, en un langage qui lui soit intelligible, des informations sur les stimulations qu’ils captent.
Le schéma de la figure 2 symbolise les trois étapes essentielles du dialogue organisme-milieu : ébranlement du récepteur, activité du système nerveux central, activité de l’effecteur.
• L’ébranlement du récepteur constitue la stimulation ; son résultat, son effet sur le récepteur, est qualifié d’excitation ; ce qui, en conséquence de l’excitation, chemine le long des voies afférentes et atteint le système nerveux central est qualifié de message sensoriel. Par exemple, la stimulation est un choc, l’excitation une perméabilité accrue du récepteur aux ions Na+, le message sensoriel un train d’ondes de dépolarisation. On voit combien il est important de ne pas employer ces trois termes l’un pour l’autre.
• L’activité du système nerveux central revêt de nombreux aspects, dont nous ne retiendrons que les plus simples.
— Les centres envoient à des effecteurs déterminés (muscles, glandes) des messages analogues par leur nature aux messages sensoriels, mais cheminant le long des voies efférentes et comparables à des « ordres » dans la mesure où ils mettent les effecteurs en action (la Grenouille retire sa patte de l’acide, etc.) [liaison 1].
— Ils reçoivent de ces mêmes effecteurs des informations en retour qui leur permettent de contrôler la parfaite exécution de leurs ordres et, s’il y a lieu, de corriger leurs instructions (liaison 2). De telles rectifications au cours de la réponse se produisent constamment dans l’équilibration, le grimper, la lutte, la manipulation de lourdes masses, etc.
— Ils améliorent éventuellement leurs informations sur le stimulus par un réglage en retour des récepteurs : les pupilles s’ouvrent pour mieux voir en faible lumière, l’oreille se rend attentive à une bande déterminée de fréquences sonores (liaison 3).
— Enfin, ils enregistrent l’événement sensoriel dans une mémoire, au moins à court terme, sans laquelle l’établissement des réflexes conditionnés serait inexplicable.
• L’activité de l’effecteur peut avoir pour résultat de faire cesser la stimulation, tenue pour désagréable (nociceptrice) : on ferme les yeux devant une trop vive lumière. Mais le processus est parfois moins direct : une piqûre de Moustique ne provoque pas, à elle seule, la vaso-dilatation locale, qui diluerait la salive irritante de l’Insecte dans une grande quantité de sang et mettrait fin à la démangeaison ; mais elle provoque un réflexe de grattage, et le grattage, à son tour, provoque la vaso-dilatation libératrice. L’effecteur a joué le rôle d’un simple relais.
Transduction et codage
La transduction est le passage de la stimulation à l’excitation. Le codage est le passage de l’excitation au message sensoriel. Ces deux transformations sont, selon les cas, réalisées par une seule cellule, par deux cellules ou par trois cellules (fig. 3).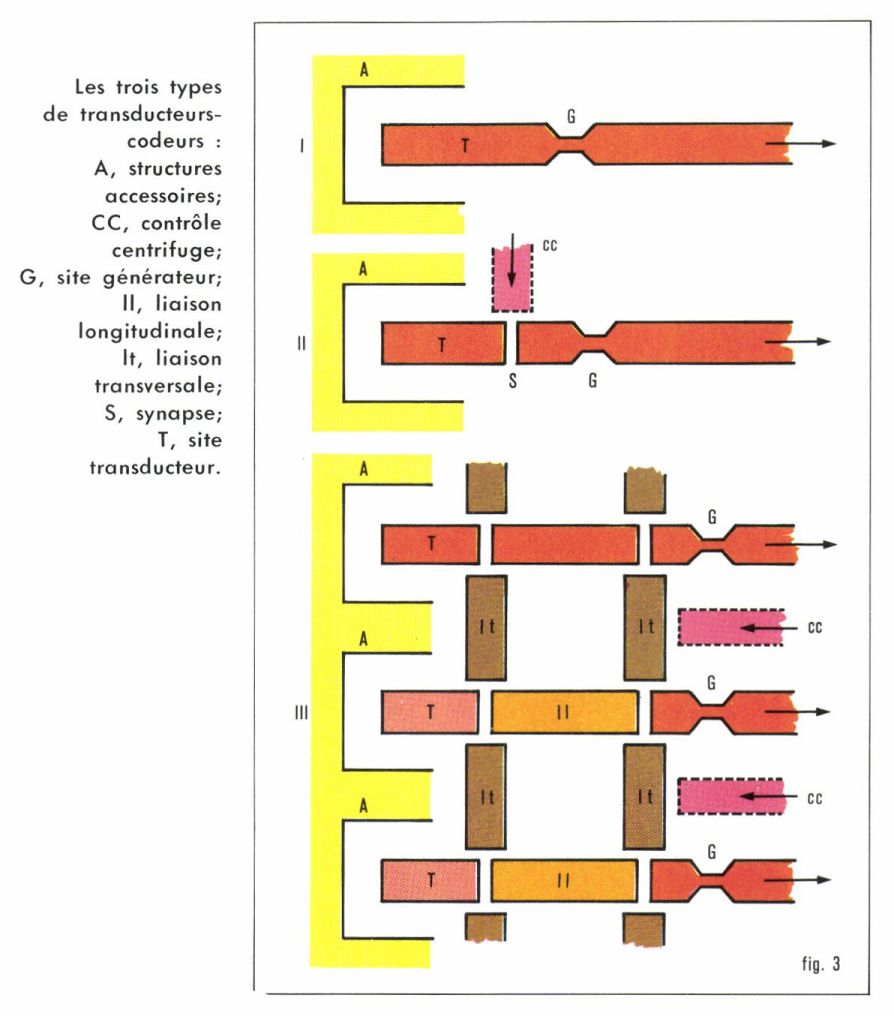
• Transducteurs-codeurs à une seule cellule (type I). Ce sont de beaucoup les plus répandus dans le monde animal, presque les seuls chez les Invertébrés ; chez les Vertébrés, ils constituent les récepteurs olfactifs, les mécanorécepteurs de la peau et les propriocepteurs musculo-tendineux. La partie distale de la cellule est un capteur-transducteur, souvent éloigné du corps cellulaire, dont le rôle est purement trophique (que l’on songe aux ganglions spinaux des racines afférentes de la moelle épinière, parfois distants de plus de 1 m du récepteur cutané !). La partie proximale est un axone.
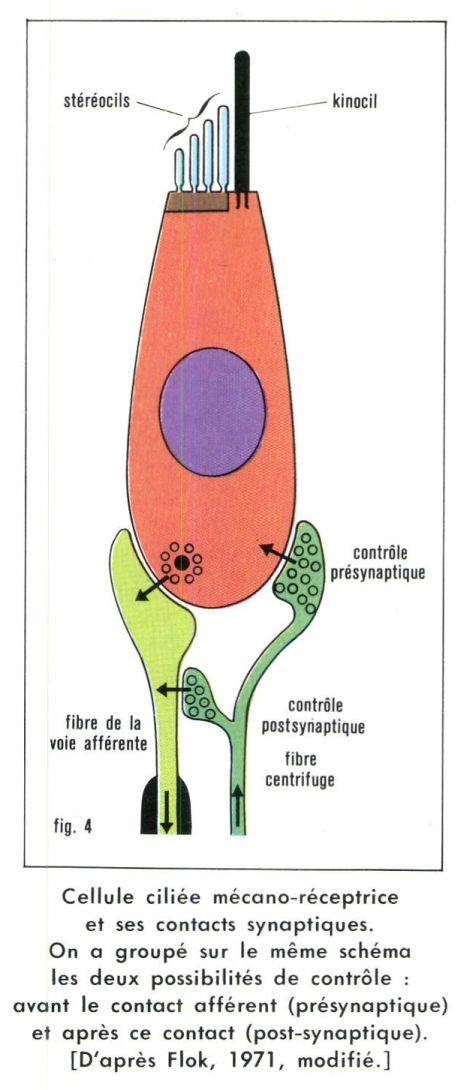
• Transducteurs-codeurs à deux cellules (type II). On y distingue une ou plusieurs cellules réceptrices, assurant seulement la transduction, et un neurone sensoriel codeur en synapse avec le récepteur. Les récepteurs du goût, de l’audition, de l’orientation spatiale (baresthésie) des Vertébrés aquatiques et aériens en sont de bons exemples. La figure 4 nous montre que, dans un tel système, un contrôle de la sensibilité par le système nerveux central peut s’exercer à deux niveaux, en amont et en aval de la synapse récepteur-neurone.