




sensation (suite)
• Transducteurs-codeurs à trois cellules (type III). Le seul exemple connu est la rétine des Vertébrés. Ici, entre la cellule réceptrice et le neurone afférent au système nerveux central, un élément de liaison (neurone bipolaire) s’intercale, permettant notamment les nombreuses liaisons transversales qui assurent, dès le niveau de l’organe sensoriel, une certaine structuration du champ et du message qui en provient. Il ne serait pas excessif de parler ici d’un prétraitement de l’information.
Cils et villosités
La microscopie électronique a permis, depuis quelques années, de s’assurer de la nature ciliaire de l’appareil transducteur des cellules réceptrices (fig. 5). Généralement, les cils ont perdu leur mobilité. Toutefois, les cellules gustatives des Vertébrés, au lieu de cils, présentent des microvillosités, ayant d’ailleurs la même fonction : augmenter énormément la surface cellulaire. Cette prédominance des structures ciliaires, surtout dans les photorécepteurs, se retrouve chez les Épineuriens et les Épithélioneuriens (Vertébrés, Procordés, Échinodermes), et même chez les Cœlentérés, tandis que les Hyponeuriens (Annélides, Mollusques, Arthropodes) présentent souvent des récepteurs non ciliés, avec empilement de lamelles ou nombreuses villosités.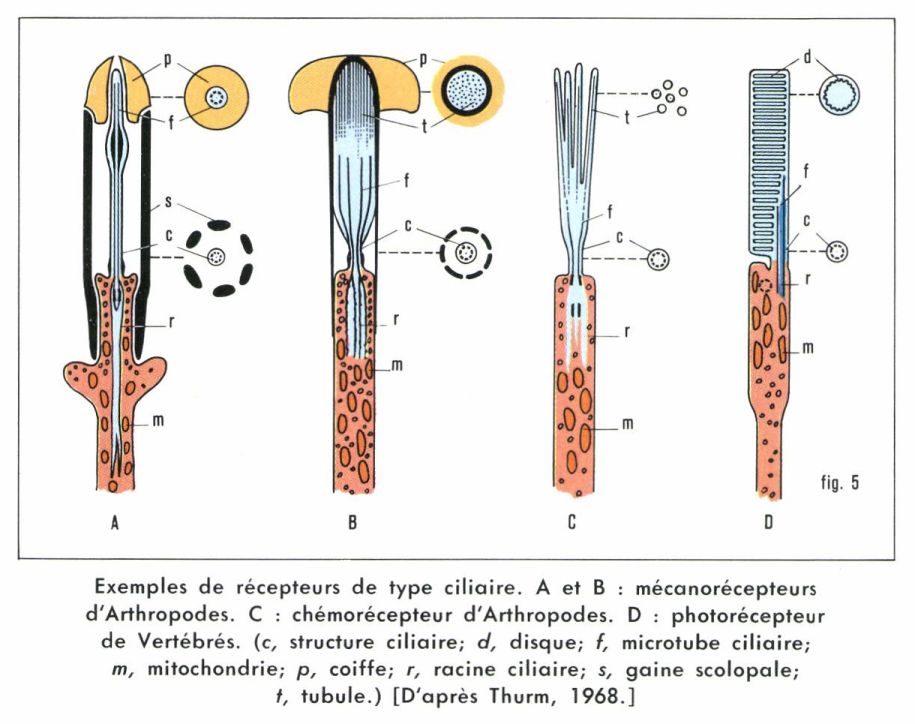
Le mécanisme de la transduction
Malgré la diversité de leur action immédiate — déformation mécanique de la membrane cellulaire (tact), apport de molécules (odorat), pénétration de photons (vision) —, les stimuli semblent avoir tous le même effet : augmenter la perméabilité de la membrane cellulaire aux ions Na+. Ceux-ci, étant au repos beaucoup plus concentrés au-dehors qu’au-dedans, envahissent alors le récepteur et le privent temporairement de sa polarité négative (dépolarisation). Ainsi se crée un potentiel de récepteur, qui décroît d’ailleurs rapidement, et toujours selon la même courbe, en fonction du temps (fig. 6). Il n’aurait pu se former si, en l’absence de toute stimulation, la concentration en ions Na+ était la même sur les deux faces de la membrane cellulaire. Or, le maintien d’une inégalité permanente de concentration se solde par une dépense d’énergie permanente, elle aussi.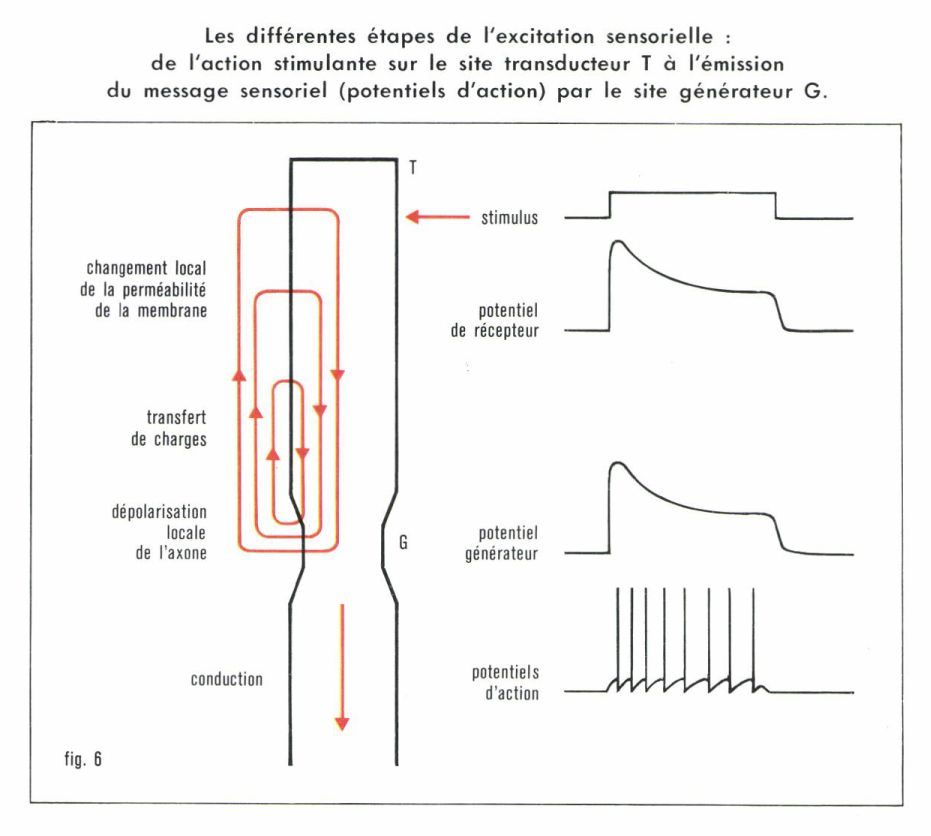
Un transducteur sensoriel n’est donc nullement un transformateur d’énergie ; c’est, au sens de la technologie moderne, un capteur actif, c’est-à-dire disposant de ses propres ressources énergétiques. On doit, ici, rendre hommage à Mach, qui, dès 1905, écrivait : « Les organes des sens sont en général très sensibles, ce qui tient à ce qu’ils ne reçoivent pas les excitations physiques passivement ; ces excitations mettent en liberté des énergies accumulées dans ces organes, chose qui n’arrive qu’exceptionnellement dans certains appareils de physique, microphone, relais télégraphique, etc. » On opposera cette formulation, en avance de près de trois quarts de siècle sur sa démonstration expérimentale, à celle de Lénine, qui est rappelée plus haut et qui ne faisait que reprendre la conception fausse d’une transformation énergétique. La stimulation libère une énergie potentielle ; elle ne la crée pas, pas plus que l’ouverture d’une vanne ne crée l’énergie du fluide libéré par cette ouverture. De même que l’énergie libérée est d’autant plus grande que la vanne est plus ouverte, de même l’amplitude du potentiel de récepteur est d’autant plus grande que l’action du stimulus sur la membrane est plus importante. On envisagera plus bas la forme de la fonction qui relie l’une à l’autre ces deux variables.
Les mécanismes du codage
Au repos, la cellule réceptrice est tout entière, d’un bout à l’autre, au même potentiel. La dépolarisation du site distal T (fig. 6) ne tarde pas à se transmettre au site proximal G, créant un potentiel générateur. Au-dessous d’une certaine valeur critique, rien d’autre ne se passe. Si, en revanche, cette valeur critique est atteinte ou dépassée, des potentiels d’action naîtront, constituant le message sensoriel proprement dit, et seront envoyés au système nerveux central, avec ou sans synapses intercalaires selon le type de récepteur considéré. Mais le message sensoriel issu d’une ou de plusieurs cellules réceptrices et transmis par une ou plusieurs fibres nerveuses est-il toujours le même lorsqu’il est issu des mêmes cellules ? Nullement. Le récepteur peut coder l’intensité de la stimulation, sa nature qualitative ou ses caractères spatio-temporels.
• Codage de l’intensité. Le premier, Gustav Theodor Fechner, dans ses Elemente der Psychophysik (1860), a insisté sur l’importance capitale de la membrane du transducteur. C’est là, en effet, que gît la discontinuité fondamentale : extérieurement à cette interface, phénomènes physiques ou chimiques ; dans la cellule, phénomènes proprement physiologiques ; au-dehors, stimulation plus ou moins intense ; au-dedans, potentiel de récepteur de plus ou moins grande amplitude. Mais quelle est la relation numérique entre ces deux grandeurs ?
• Lorsqu’on porte en abscisses le logarithme de l’intensité stimulatrice et en ordonnées l’amplitude du potentiel de récepteur, on obtient une courbe du type de celle qui est représentée à la figure 7, et cela dans les expériences les plus diverses, qu’il s’agisse de mécanorécepteurs, de photorécepteurs ou de chémorécepteurs. Les incurvations de la courbe sont telles que la simple relation logarithmique fechnérienne n’est valable que dans la marge étroite des valeurs moyennes, mais on peut écrire la fonction qui rend compte de la réalité expérimentale et qui est du type « tangente hyperbolique ».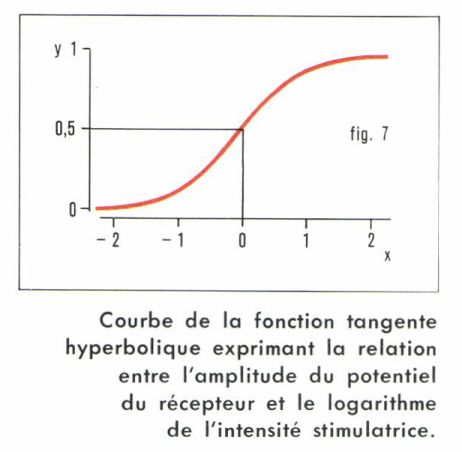
Sans reproduire ici le détail des calculs, mentionnons simplement la formule sur laquelle on s’accorde :
Dans cette formule,  V max étant le plus grand potentiel que le récepteur soit capable de fournir et V le potentiel qu’il fournit en réponse au stimulus d’intensité I ;
V max étant le plus grand potentiel que le récepteur soit capable de fournir et V le potentiel qu’il fournit en réponse au stimulus d’intensité I ;  étant une fonction linéaire assez simple de I. Celte courbe présente un centre de symétrie pour
étant une fonction linéaire assez simple de I. Celte courbe présente un centre de symétrie pour  (c’est-à-dire pour thx = 0).
(c’est-à-dire pour thx = 0).