




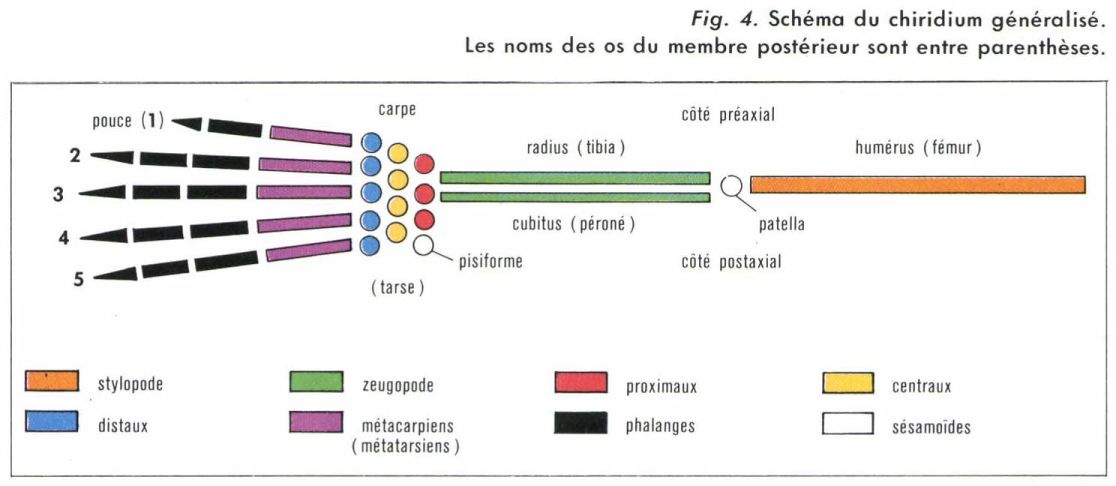
membres (suite)
Chez les Tétrapodes, on distingue deux types de membres : le transversal et le parasagittal. Dans le membre transversal, le plus primitif (Batraciens, la plupart des Reptiles), le stylopode est perpendiculaire à l’axe du corps, le zeugopode lui étant nécessairement orthogonal. L’effort de sustentation est alors considérable, ce qui réduit la puissance disponible pour la marche. Dans le membre parasagittal (Oiseaux, Mammifères), stylopode et zeugopode sont tous deux situés dans un plan parallèle au plan de symétrie du corps. L’effort de sustentation est alors réduit au minimum ; de plus, cette disposition autorise l’allongement du membre et lui confère une grande souplesse.
• Quelques variations adaptatives.
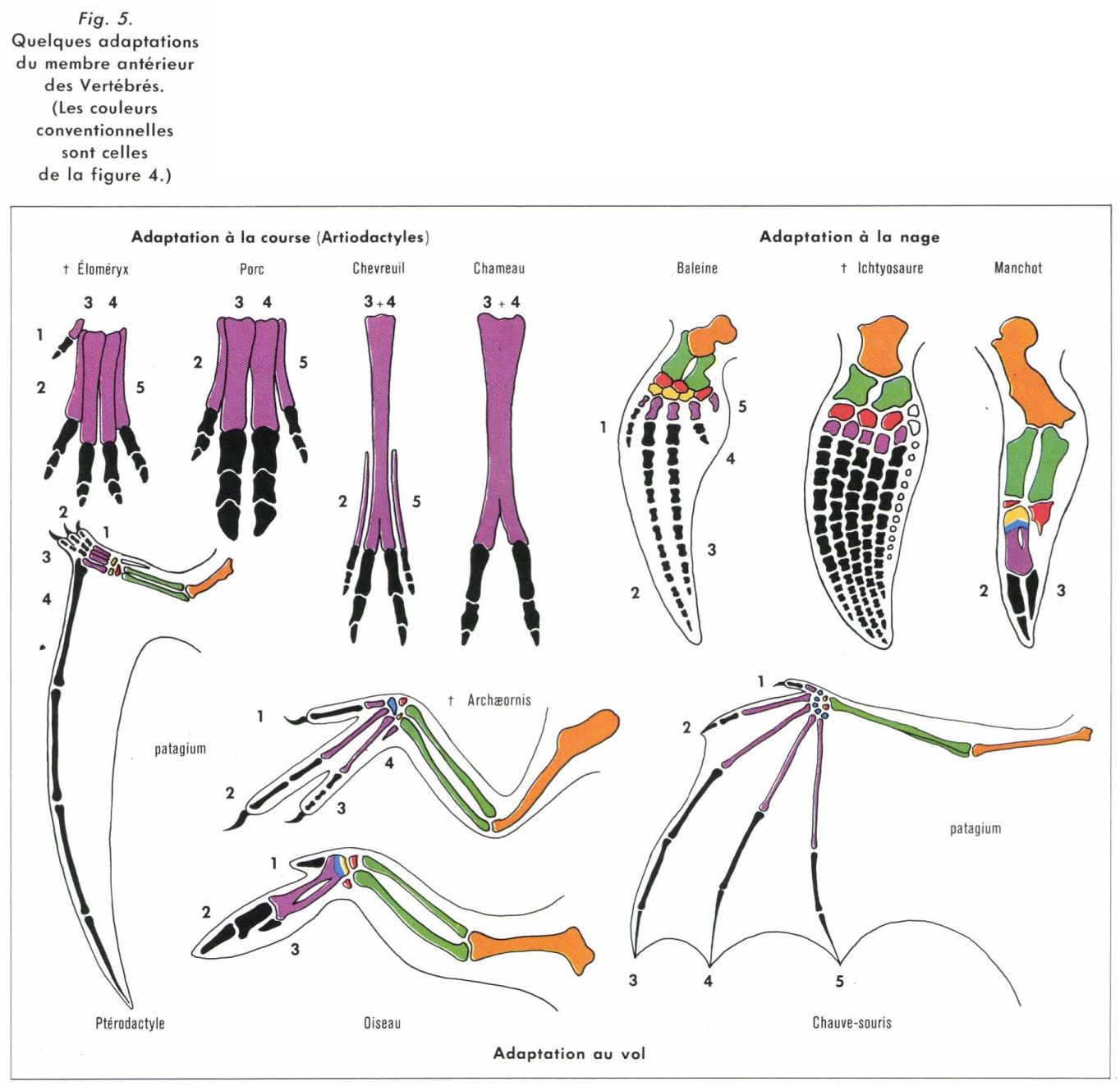
Adaptation à la course. Particulièrement spectaculaire chez les Mammifères, elle se manifeste par la réduction du nombre des doigts et une tendance vers le type onguligrade (réduction du contact avec le sol) ainsi que par l’allongement des métacarpiens et des métatarsiens (augmentation de l’amplitude des mouvements). Chez les Ongulés, on distingue une série artiodactyle : prédominance des doigts 3 et 4 (qui subsistent seuls chez le Bœuf et le Chameau) avec fusion des métacarpiens ou métatarsiens correspondants en un os canon, et une série périssodactyle : prédominance du doigt 3, qui subsiste seul chez le Cheval.
Adaptation au vol. Chez les Oiseaux, elle se manifeste notamment par la réduction du nombre des osselets libres du poignet, avec fusion des centraux et des distaux entre eux et aux métacarpiens (carpométacarpe), et par la réduction du nombre des doigts et des phalanges. De grandes plumes, les rémiges, s’insèrent sur toute la longueur du bras, auquel elles fournissent une grande surface portante. Cette évolution était déjà bien amorcée chez Archæornis et Archæopteryx, qui possédaient cependant encore trois doigts griffus et libres.
Chez les Chauves-Souris, le cubitus est réduit (comme il est fréquent chez les Mammifères), les centraux ont disparu, les autres carpiens sont libres. Les cinq doigts subsistent et, sauf pour le premier, qui est libre, les métacarpiens et les phalanges (dont le nombre est variable) sont très allongés et soutiennent la membrane patagiale qui assure la surface portante nécessaire. Chez les Ptérodactyles (Reptiles volants fossiles), il n’y a que quatre doigts, le quatrième, démesuré, soutient seul le patagium.
Adaptation à la nage. Chez les Tétrapodes qui ont fait retour à la vie aquatique, notamment certains Reptiles (Ichtyosaures, Tortues de mer...) et Mammifères (Phoques, Cétacés...), les membres sont transformés en nageoires, mais sans retour à la structure ptérygienne : la structure chiridienne est conservée. Il s’agit d’une convergence. Les modifications sont le raccourcissement des os du stylopode et du zeugopode (généralement aplatis), la réduction du nombre des carpiens (et des tarsiens lorsque le membre postérieur est conservé) et la perte de la liberté des doigts. Ceux-ci, dont les phalanges sont souvent très nombreuses (hyperphalangie), sont réunis par du conjonctif recouvert par la peau ; le membre prend alors l’aspect d’une pagaie. La transformation en nageoire peut affecter un membre déjà spécialisé, qui conserve alors sa structure caractéristique tout en subissant la nouvelle variation adaptative : ainsi l’aile du Manchot, dont les os sont courts et plats, ne sert qu’à la nage et est dépourvue de rémiges.
Notons enfin que les membres peuvent disparaître chez les formes allongées (Serpents, certains Lézards et Batraciens). Toutefois, les Boas conservent des rudiments de membres postérieurs, dont seul le tibia, en forme de griffe, est décelable de l’extérieur.
• Origine du chiridium.
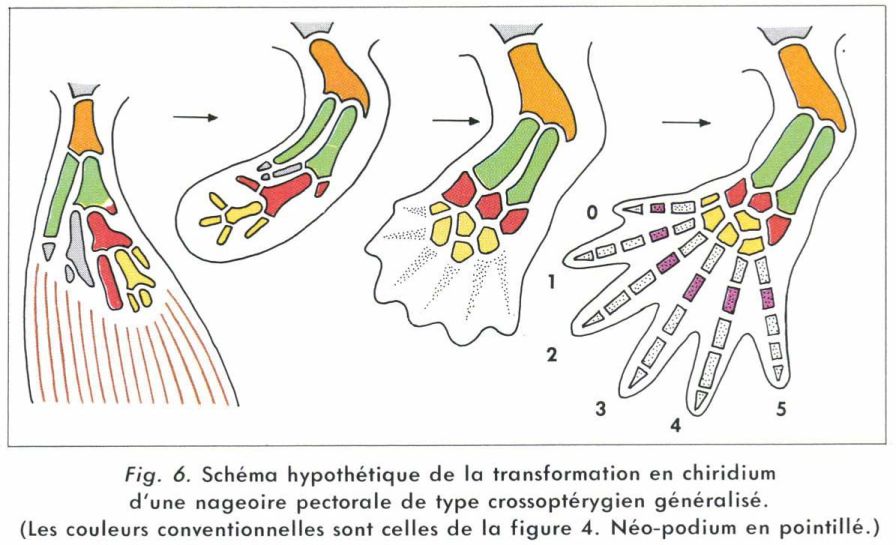
La ressemblance entre les squelettes fossiles de la patte des Labyrinthodontes primitifs (Batraciens) et de la nageoire de certains Crossoptérygiens Rhipidistides conduit à imaginer l’origine de celle-là à partir de celle-ci. Toutefois, les carpiens (et tarsiens) distaux, les métacarpiens (et métatarsiens) et les phalanges seraient des néo-formations (néo-podium). Il est en outre probable que la patte primitive comportait un doigt supplémentaire en avant du pouce (noté 0). La figure 6 schématise l’une des nombreuses théories émises à ce sujet.
Les membres de l’homme
L’histoire des membres humains commence à cette lointaine époque où le premier amphibie émergea de la surface des eaux : ses membres comportaient déjà un segment proximal, un segment intermédiaire et un segment distal muni de cinq doigts. Au Miocène, il y a quelque vingt millions d’années, c’est le pied* d’un petit animal arboricole de la famille des Lémuriens qui, plus que le pied du singe, se rapproche du nôtre. Le pied préhistorique, avec son gros orteil dirigé en dedans, nettement séparé des autres, était souple, préhensile, mais plat. La coupole plantaire n’apparut que plus tard, quand l’homme adopta la station verticale. Ce passage à l’Homo erectus, événement considérable, modifia notre statique entière : pour maintenir en équilibre tête, rachis, hanche, genou et tibio-tarsienne, il fallut une vigilance musculaire constante. Tandis que la main se spécialisait dans la préhension fine, le pied s’adapta à la fonction portante.
Embryogenèse des membres
Chez l’embryon, les membres commencent à apparaître vers la troisième semaine, sous forme de simples palettes qui s’accroissent rapidement, se divisent en deux puis trois articles ; sur le troisième article apparaissent quatre sillons qui, en s’accentuant, sépareront progressivement les ébauches des doigts ou des orteils. Les membres sont d’abord perpendiculaires à l’axe de l’embryon ; ils prennent ultérieurement leur position définitive par une rotation de 90° en sens inverse : le membre supérieur fait une rotation externe qui dirige son articulation en avant, le coude en arrière ; le membre inférieur exécute une rotation interne qui amène son articulation en arrière et le genou en avant.