




développement et différenciation (suite)
La gastrulation
Cette étape, la plus importante peut-être du développement embryonnaire des Métazoaires, correspond à l’ensemble des processus morphogénétiques mettant en place les feuillets fondamentaux : deux chez les organismes diploblastiques (Spongiaires, Cnidaires), trois chez les autres Métazoaires (triploblastiques). Bien que les mitoses n’aient pas cessé, mais soient simplement devenues moins fréquentes, ce sont surtout les migrations cellulaires qui caractérisent la gastrulation. Cette fois, les modalités en s’ont plus en rapport avec la répartition systématique des espèces étudiées qu’avec la structure de la cellule initiale. Malheureusement, l’observation banale ne rend que très mal compte de ces phénomènes. L’analyse n’en a été rendue possible que grâce à la technique des marques colorées, mise au point par Walther Vogt (né en 1918) : l’œuf d’Amphibien, débarrassé de sa gangue, est localement teinté par des colorants vitaux (rouge neutre, bleu de Nil). Le colorant diffuse et imprègne certaines cellules, qui restent colorées pendant plusieurs jours et qu’on peut retrouver à l’intérieur de l’embryon si elles y ont émigré. En repérant, après coup, l’ensemble des régions dont on a suivi l’évolution sur un modèle de blastula, on établit une carte des territoires présomptifs (fig. 2).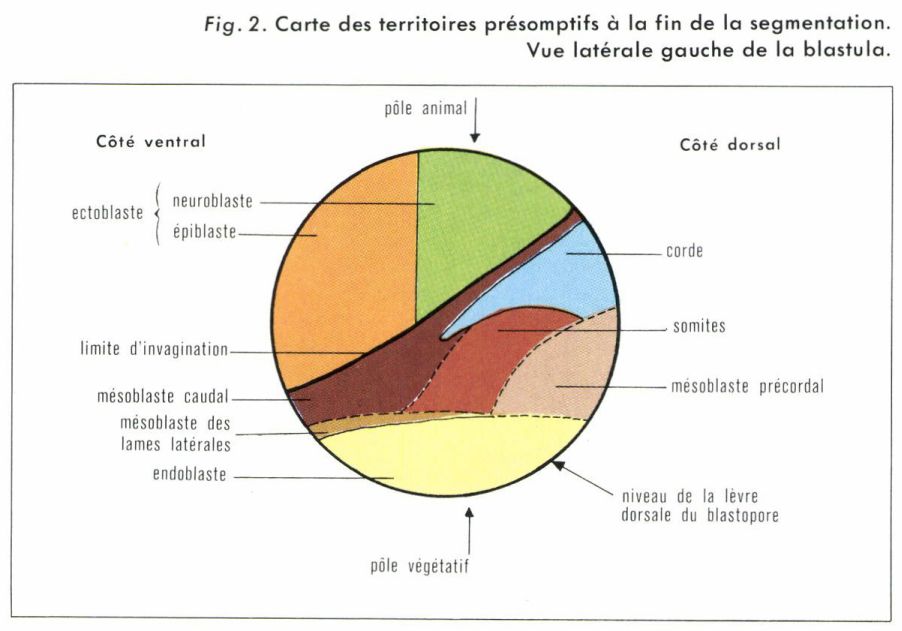
Du côté dorsal, juste sous le croissant gris maintenant estompé, se forme une encoche, ou blastopore, dont le rôle va se révéler fondamental. La lèvre dorsale du blastopore s’étend latéralement ; la région ventrale s’enroule en profondeur et s’invagine, des cellules jusque-là externes passant en position interne ; on parle de gastrulation par embolie. Le feuillet interne, ou endoblaste, délimite l’intestin primitif, ou archentéron. Au fur et à mesure que celui-ci s’agrandit et progresse, le blastocèle se réduit et disparaît. Du côté dorsal, la lèvre du blastopore s’enroule et invagine du matériel cordomésoblastique. À la fin du processus, les lèvres blastoporales se joignent ventralement et entourent la zone dépigmentée inférieure de l’embryon, qui forme le « bouchon vitellin ». Celui-ci s’invagine jusqu’à disparaître à peu près totalement. L’hémisphère supérieur de la blastula (ectoblaste présomptif) recouvre les autres feuillets en un mouvement qualifié d’épibolie.
En définitive, il y a invagination, embolie de l’endoblaste et du cordo-mésoblaste, alors que l’ectoblaste évolue par épibolie (fig. 3). L’ensemble du processus dure vingt-quatre heures chez les Amphibiens. Avec des modalités variables et un résultat qui dépend du plan d’organisation dans le groupe étudié, les deux ou trois feuillets, selon le cas, sont mis en place à la fin de la gastrulation, mais cela ne signifie pas que les mouvements morphogénétiques soient terminés. Au cours de la période suivante, des mouvements vont encore s’observer, la forme de l’embryon s’ébauchant alors, en même temps que s’accentue l’utilisation par l’embryon des réserves qui l’accompagnent, ce qui permet la croissance, jusque-là à peu près nulle.
Organogenèse
Les mouvements varient beaucoup selon les groupes zoologiques, mais c’est pendant cette période que les cellules acquièrent leur détermination définitive et qu’elles se différencient pour former les tissus.
Chez les Amphibiens et plus généralement chez les Cordés, un épaississement de la région dorsale, au-dessus de la fente blastoporale, forme la plaque neurale. L’embryon perd sa forme sphérique et s’allonge dans un sens antéropostérieur : c’est le stade neurula (fig. 4). Les bords épaissis de la plaque neurale se soulèvent, vont à la rencontre l’un de l’autre pour former le tube nerveux, pendant que le mésoblaste dorsal s’épaissit et se creuse d’une cavité, ou « cœlome ». Le tube nerveux s’enfonce. Sa région antérieure, plus dilatée, sera à l’origine de l’encéphale. Le mésoblaste dorsal se découpe en blocs réguliers, ou somites, à partir desquels se différencieront les éléments du squelette axial et de la musculature dorsale. Les territoires présomptifs sont ainsi irrémédiablement engagés quant à leur devenir : ils sont déterminés. C’est seulement lorsque leur mise en place sera achevée qu’ils se différencieront histologiquement. La complexité des processus de mise en place des organes est telle qu’il n’est pas possible de les décrire ici, bien qu’ils soient parfaitement connus. Lorsque les organes sont devenus fonctionnels, l’embryon est capable de mener une vie libre : le développement embryonnaire est terminé. Chez la Grenouille, par exemple, l’embryon est parvenu au stade du bourgeon caudal (fig. 5), très semblable au têtard, libéré par l’éclosion quatre jours environ après la ponte et qui mesure alors 6 mm.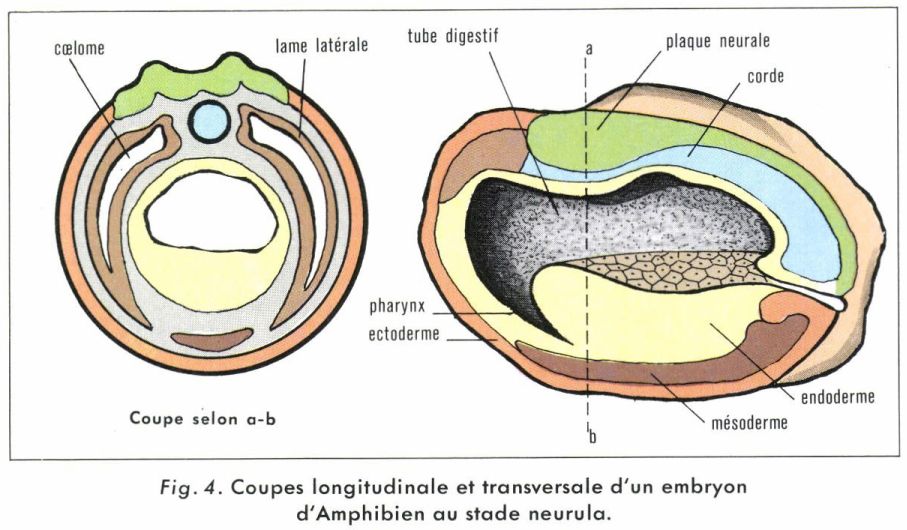

Analyse expérimentale du développement embryonnaire
La naissance de l’embryologie descriptive et les progrès que celle-ci fit au xixe s., progrès liés à l’amélioration des instruments d’observation et au développement des techniques histologiques, firent justice de la conception préformiste, selon laquelle le germe ne serait qu’une réduction de l’organisme adulte. La théorie épigénétique, formulée dès 1759 par le médecin allemand Caspar Friedrich Wolff (1733-1794), qui observa avec une précision remarquable pour l’époque la réalisation progressive de l’embryon d’Oiseau à partir de l’œuf, soutient que chaque partie de l’œuf est totipotente au début du développement et évolue peu à peu, formant un organe de l’embryon, puis de l’organisme définitif. Mais il est bientôt apparu que chaque territoire embryonnaire perdait sa totipotence pour acquérir un destin organogénétique déterminé et limité bien avant que les effets de cette détermination fussent visibles : il y a une sorte de prédifférenciation.
La détermination des territoires de l’embryon
Tout œuf manifeste à des degrés divers une certaine tendance à l’anisotropie : la répartition des réserves et des ribonucléoprotéines, par exemple, s’effectue selon deux gradients de direction opposée, le vitellus s’accumulant du côté du pôle végétatif, alors que les ribonucléoprotéines sont plus abondantes vers le pôle animal. Cette anisotropie est telle dans certains groupes (Mollusques, Annélides, Ascidies) que l’ablation d’un blastomère à un stade très précoce de la segmentation s’accompagne d’une perte définitive de tout ou partie d’un organe, toujours le même pour une même cellule enlevée : on parle alors d’œuf à développement mosaïque.