




parasitisme (suite)
Adaptations morphologiques des parasites
La vie parasitaire a entraîné des adaptations morphologiques très particulières, parmi lesquelles il faut notamment signaler le dimorphisme* sexuel, la forme aplatie du corps, les organes de fixation et l’absence de tube digestif.
Dimorphisme sexuel
Chez certains parasites, le mâle est minuscule par rapport à la femelle. On observe ce fait chez de nombreux Crustacés (Copépodes, Isopodes, Cirripèdes), chez des Vers (Myzostomides, Échiurides, Nématodes). Parfois, le mâle est lui-même parasite de la femelle ; tel est le cas chez beaucoup de Crustacés et chez des Vers (ainsi, le mâle du Nématode Trichosomoides, parasite de la vessie du Rat, vit fixé dans l’utérus de la femelle). Il y a ici hyperparasitisme (parasitisme d’un organisme lui-même parasite).
Dans d’autres cas, c’est la femelle qui est beaucoup plus petite que le mâle et qui est logée dans une rainure de son corps (par exemple le Trématode du genre Bilharzia).
Chez certains parasites (Crustacés, Insectes, Nématodes), les femelles perdent leurs appendices et deviennent des sacs remplis d’œufs avec une fréquente hypertrophie de l’appareil génital (fig. 2). Nous reviendrons plus loin sur cette fécondité accrue.
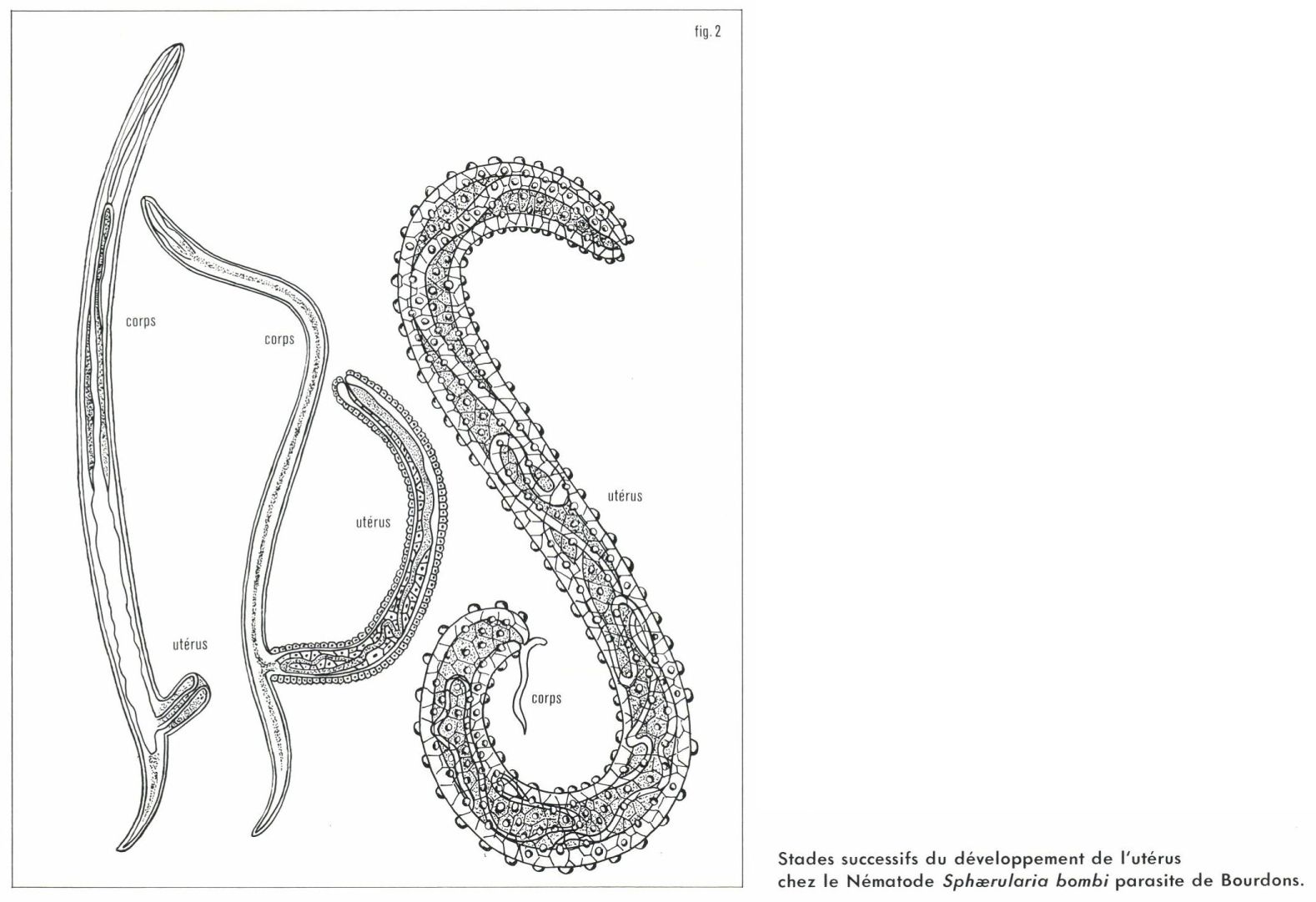
Forme aplatie du corps
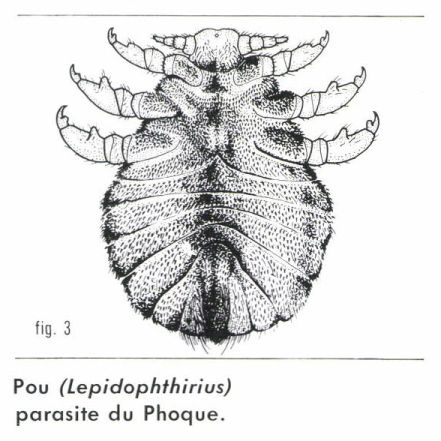
C’est le cas chez beaucoup d’Insectes ectoparasites de Vertébrés à sang chaud, qui peuvent ainsi circuler plus facilement dans la fourrure ou le plumage de leur hôte. L’aplatissement peut être dorso-ventral (Poux [fig. 3], Punaises, divers Acariens) ou latéral (Puces). Chez certains d’entre eux, on observe d’autres adaptations encore plus spécialisées. Ainsi, chez un Pou (Lepidophthirius) parasite du Phoque, le corps est recouvert de minuscules écailles qui retiennent l’air lorsque l’hôte plonge.
Organes de fixation
Ceux-ci sont de nature très diverse (griffes, crochets, ventouses, mucrons, suçoirs, etc.) et se rencontrent dans les groupes les plus divers de parasites, dont ils assurent la fixation sur l’hôte. On les observe aussi bien chez des Protozoaires fixés à l’épithélium intestinal de l’hôte (Grégarines) que chez des helminthes (ventouses et crochets des Trématodes et des Cestodes). Chez certains Ténias, c’est toute l’extrémité antérieure (scolex) qui se fixe dans la paroi intestinale de l’hôte en réalisant un dispositif analogue à celui du bouton-pression.
Beaucoup d’Insectes ectoparasites ont des appendices munis de griffes, de crochets ou d’épines qui leur permettent de s’accrocher aux poils et aux plumes de l’hôte. Les cténidies, rangées d’épines serrées l’une contre l’autre et rappelant les dents d’un peigne, se rencontrent chez divers Insectes ectoparasites de Mammifères (Puces, Poux, Punaises, Coléoptères, Diptères) et ne se trouvent jamais chez des espèces libres.
On note également la présence de griffes, de pinces ou de stylets perforants chez les Crustacés parasites.
Absence de tube digestif (agastrie)
Celle-ci s’observe chez certains Copépodes (Monstrillides, Lamippides), et c’est la règle absolue chez les Cestodes. Cette agastrie est compensée par la présence d’organes absorbants tels que les rhizoïdes de divers Copépodes. Chez les Cestodes, la nutrition se fait par osmose à travers le tégument, dont les très nombreuses microvillosités augmentent la surface d’absorption (il en est de même chez divers Protozoaires parasites et symbiotiques).
Adaptations biologiques et biochimiques des parasites
Les principales adaptations biologiques des parasites sont liées à leur sexualité et à leur reproduction. Pour beaucoup d’endoparasites, la rencontre des sexes est rendue difficile par leur localisation dans la profondeur des organes ou des tissus de l’hôte. Cet inconvénient est pallié de diverses façons : tout d’abord par l’hermaphrodisme, avec possibilité d’autofécondation (Trématodes, Cestodes, Rhizocéphales, certains Isopodes et Nématodes). Chez d’autres parasites, les deux sexes vivent réunis. C’est le cas des Bilharzia (Trématodes), de la plupart des Épicarides (Isopodes) et de presque tous les Copépodes.
Par ailleurs, le cycle souvent compliqué des parasites, nécessitant la présence d’un ou de plusieurs hôtes intermédiaires avant la rencontre avec l’hôte définitif (voir le paragraphe qui suit), et l’existence de migrations amènent la perte d’un grand nombre d’embryons ou de larves. Cela pourrait entraîner la disparition de l’espèce si celle-ci ne possédait pas une très grande fécondité, souvent en relation avec l’hypertrophie des femelles déjà mentionnée. En voici quelques exemples précis : la cavité incubatrice d’un Épicaride renferme plusieurs milliers d’embryons ; l’Ascaris (ou Ascaride) de l’Homme (Ascaris lumbricoides) pond annuellement 64 millions d’œufs ce qui représente 1 700 fois son poids ; l’Ankylostome (Ankylostoma duodenale) en pond de 5 000 à 10 000 par jour et peut vivre de quatre à cinq ans, ce qui fait un total de plus de 18 millions d’œufs pondus pour une seule femelle ; le Ver solitaire (Tænia solium) en émet 80 millions par an, et le Bothriocéphale (Diphyllobothrium latum), qui peut vivre plus de dix ans, produit un nombre de segments pouvant atteindre une longueur de 7 km et contenant environ 2 billions d’œufs ; la Douve du foie (Fasciola hepatica) peut pondre théoriquement 37 000 œufs donnant plus de 6 millions de cercaires.
D’autres processus biologiques favorisent la multiplication des parasites : la schizogonie, reproduction asexuée qui s’observe chez divers Protozoaires (Grégarines, Coccidies) ; la strobilisation, ou bourgeonnement des Cestodes larvaires (cysticerques) ; la différenciation des rédies dans les sporocystes des Trématodes digénétiques, donnant elles-mêmes de nombreuses cercaires ; les phénomènes de bourgeonnement chez certains Cirripèdes (Rhizocéphales) ; la polyembryonie (formation de plusieurs embryons à partir d’un seul œuf) chez les Insectes entomophages (Hyménoptères chalcidiens et Proctotrypides).