L’Essai sur l’entendement humain (Essay concerning human understanding), qui est l’ouvrage capital de Locke, se compose de quatre livres. Le premier réfute la théorie des idées innées ; le second décrit la genèse des « idées de sensation » à partir des sens externes et des « idées de réflexion » à partir du sens interne ; le troisième, celle des idées générales par l’intermédiaire du langage ; le dernier fixe ce qu’il est permis, étant donné l’origine et la nature de ces idées, d’exiger et d’espérer de l’esprit humain tant en matière scientifique que religieuse.
D. H.
➙ Lumières (Esprit des).
N. Reicyn, la Pédagogie de John Locke (Hermann, 1941). / R. Polin, la Politique morale de John Locke (P. U. F., 1960).
locomotion
Action de changer de lieu par déplacement actif autonome.
Il convient d’éliminer de la notion de locomotion tout transport passif : la Physalie, dont le flotteur plein de gaz fonctionne comme une voile que pousse la brise du large, la jeune Araignée suspendue à un long fil de soie (fil de la Vierge) que promène le vent ne sont que des voyageurs passifs.
Les mouvements complexes du déplacement actif nécessitent un ensemble coordonné de réflexes et un point d’appui contre lequel puisse s’exercer une poussée. Dans l’immense majorité des cas, cette poussée, exactement égale et opposée à la force propulsive, s’exerce sur un point localisé du milieu. Il est évident que la localisation de ce point d’appui doit être temporaire par rapport à la surface du corps.
Les mécanismes de base
La propriété du cytoplasme de convertir l’énergie chimique en énergie mécanique est à la base de tout mouvement animal. Il existe trois types de tels convertisseurs : un type diffus (mouvement amiboïde), un type localisé dans des organites cellulaires (mouvements flagellaire et ciliaire) et un type localisé dans des différenciations intracellulaires : myonèmes et myofibrilles (mouvement musculaire).
Les deux premiers types sont liés aux milieux liquides.
Mouvement amiboïde
Il est caractérisé par la formation de pseudopodes, expansions temporaires d’une cellule. Ce moyen de propulsion n’est guère utilisable que par des cellules isolées : Amibes, éléments figurés du sang. Toutefois, l’Hydre peut se déplacer par glissement grâce à une action de type pseudopodial des cellules de son disque basai. (V. Amibiens.)
Mouvements flagellaire et ciliaire
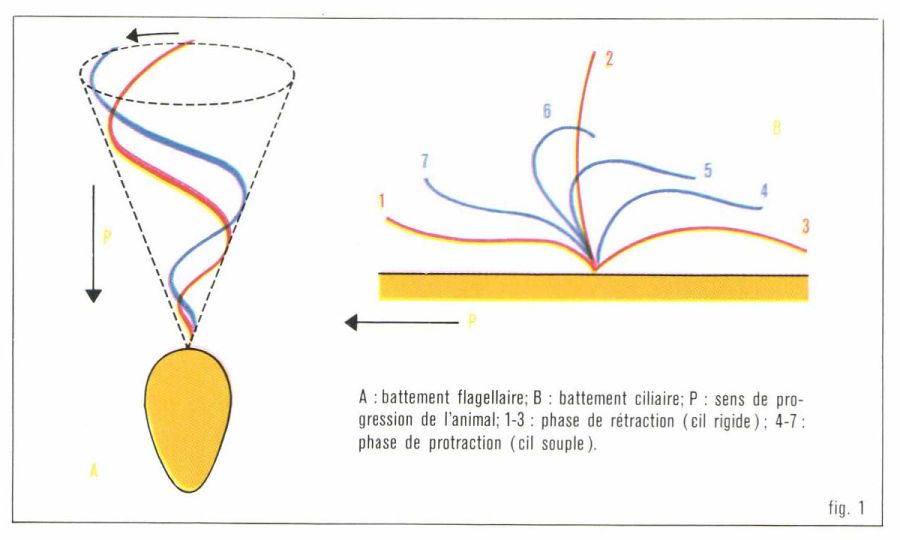
Les flagelles décrivent souvent une courbe hélicoïdale engendrant un cône de révolution : en quelque sorte, ils se vissent dans l’eau et fonctionnent comme une hélice de navire (fig. 1). Ce type de locomotion, de puissance relativement faible, ne peut être utilisé que par des cellules isolées (Protistes*, spermatozoïdes...), tout au plus par des colonies de Protistes.
Les cils, courts et plus nombreux, permettent des mouvements plus efficaces. Un cil, rigide lorsqu’il bat vers l’arrière (phase active du mouvement ou phase de rétraction), est souple lorsqu’il revient, plus lentement, vers l’avant (phase de protraction). Un déphasage régulier entre l’état cinétique des cils appartenant à deux rangées consécutives (mouvement métachrone) détermine sur la surface cellulaire des ondes qui rappellent ce que l’on voit lorsqu’un champ de céréales est caressé par le vent.
Le mouvement ciliaire est utilisé comme moyen de locomotion non seulement par des Protozoaires comme la Paramécie, mais par les larves nageuses de groupes très divers (Eponges, Cnidaires, Bryozoaires, Mollusques, Annélides, Échinodermes...), voire par des adultes (Gastrotriches).
Chez les animaux de taille plus élevée, le mouvement ciliaire est encore efficace si la viscosité du milieu est localement accrue. Les Turbellariés, les Némertes, etc., sécrètent une fine couche de mucus sur un substrat suffisamment rigide (corps solide ou film de tension superficielle) ; l’épaisse toison ciliaire de leur face ventrale peut prendre appui sur le film muqueux.
Mouvement musculaire
C’est le plus puissant et le plus efficace. Après s’être contractée, une fibre musculaire ne retrouve sa longueur initiale que si elle est étirée. Le mécanisme du mouvement musculaire requiert donc, pour la « remise à zéro » des éléments contractiles, le jeu d’un couple de systèmes antagonistes réagissant l’un sur l’autre soit par un complexe de leviers, soit par un liquide interne et rendant au muscle sa longueur initiale.
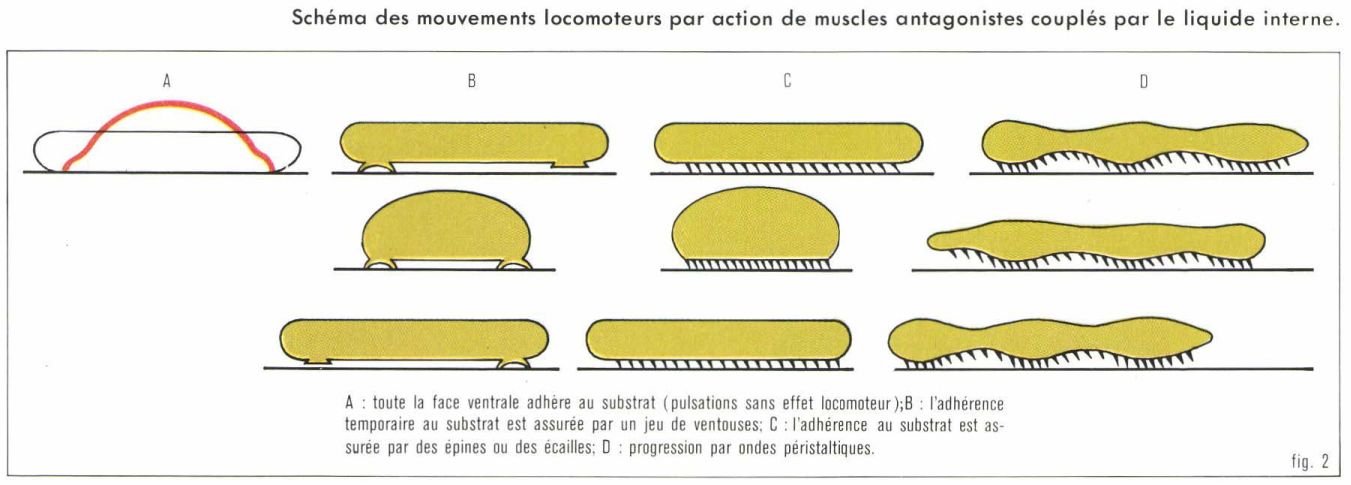
Dispositif à liquide
Imaginons un modèle d’animal doté d’une double couche musculaire pariétale circulaire et longitudinale. La contraction des muscles circulaires allonge le corps et, grâce à la présence du liquide interne dont le volume est constant, étire les muscles longitudinaux antagonistes. La contraction des muscles longitudinaux raccourcit et épaissit le corps. Ces pulsations n’ont d’effet locomoteur que si l’extrémité antérieure du corps adhère seule au substrat lors de la phase de raccourcissement et si l’extrémité postérieure du corps adhère seule lors de la phase d’élongation. L’adhérence labile peut être réalisée par du mucus, des ventouses ou des aspérités (soies, crochets, épines, etc.) qui, si elles sont dirigées vers l’arrière, fonctionnent comme cliquets (fig. 2).
Ces raccourcissements et élongations successifs peuvent intéresser seulement des portions successives du corps, ce qui fait apparaître des ondes de contraction dites « péristaltiques ». Si ces ondes se déplacent dans le sens de progression de l’animal, elles sont dites « directes » ; « rétrogrades » si elles se déplacent en sens inverse.
Si les muscles longitudinaux interviennent seuls de façon active, il apparaît des ondes sinusoïdales rétrogrades dont la figure 3 montre les conséquences mécaniques, qui se traduisent par la progression de l’animal. C’est ainsi que se déplacent les Annélides Polychètes notamment. En fait, chez ces animaux, les parapodes (expansions latérales du corps) contribuent d’une façon importante à la locomotion en fonctionnant comme pagaies. Les parapodes situés au niveau d’une convexité sont nettement séparés les uns des autres ; en se rabattant vers l’arrière, ils exercent individuellement une poussée sur le milieu. Le battement de retour s’effectue pendant que les parapodes sont situés au niveau des concavités : pressés les uns contre les autres, ils ne peuvent agir individuellement et leur action de freinage est négligeable.