Annélides (suite)
La reproduction sexuée
Chez les Polychètes, les Sipunculiens et les Échiuriens, les sexes sont séparés ; l’hermaphrodisme est la règle chez les Oligochètes, les Hirudinées et les Myzostomidés. Si les gonades individualisées existent chez les Oligochètes et les Hirudinées, chez les Polychètes les cellules germinales se forment dans la paroi des sacs cœlomiques et tombent dans la cavité générale à maturité ; dans certains cas, elles empruntent, pour sortir, les néphridies ou des pavillons génitaux ; dans d’autres, elles ne sont libérées que par rupture du corps et mort de l’animal.
La maturité sexuelle s’accompagne, chez plusieurs Polychètes, d’importantes modifications morphologiques, qui ont reçu le nom d’épitoquie, ou épigamie. Nereis en offre l’exemple le plus net : tandis que les yeux grossissent et que palpes et antennes se réduisent, la partie postérieure du corps acquiert des expansions foliacées sur chaque anneau et des soies en palettes ; le tube digestif et certains muscles subissent une histolyse ; l’animal, de rampant qu’il était, devient nageur (Heteronereis) ; lors de phases lunaires déterminées, les individus sexués montent à la surface de la mer, où a heu la fécondation. L’épitoquie se manifeste d’une manière particulière dans chaque espèce : chez Tylorhynchus chinensis, des eaux saumâtres d’Extrême-Orient, la région postérieure se détache et les éléments sexuels se développent dans la partie antérieure ; chez Syllis, c’est l’arrière qui porte les gamètes, en se séparant de l’avant et en bourgeonnant une tête, ce qui lui permet de mener une courte vie libre jusqu’à la fécondation. Eunice viridis, des îles Samoa, doit sa célébrité au fait que la montée de millions d’individus épitoques se produit à un moment précis du cycle lunaire de novembre et que les indigènes pèchent activement les Vers, qu’ils nomment « palolo ».
Ainsi rassemblés, les Polychètes sexués manifestent une activité prodigieuse, nageant en tous sens, donnant l’impression que la mer bouillonne. Les mâles se groupent autour des femelles (on parle parfois de « danse nuptiale »), mais il n’y a jamais accouplement véritable ; les cellules reproductrices sont émises dans la mer ; on a montré que les femelles produisaient une fertilisine spécifique qui provoque l’expulsion des spermatozoïdes.
Si, chez la majorité des Polychètes, le développement conduit à une larve trochophore planctonique, les embryons de certaines espèces restent fixés quelque temps à leur mère ou sont conservés dans le tube de l’adulte ou dans les chambres incubatrices.
Les Oligochètes, hermaphrodites, s’accouplent et se fécondent mutuellement, maintenus par des anneaux de mucus fourni par le clitellum. D’une famille à l’autre, la position des gonades et des orifices génitaux sur le corps varie, mais les conduits génitaux restent toujours indépendants des néphridies. Les œufs se développent dans un cocon, sans donner de larve libre.
Les Hirudinées, également hermaphrodites, s’accouplent, mais la fécondation n’est pas forcément réciproque. Chez certaines Sangsues, le « mâle » introduit ses gamètes directement dans les voies femelles. D’autres réalisent une fécondation originale, hypodermique : le mâle pique les spermatophores dans les téguments d’un individu fonctionnellement femelle ; les spermatozoïdes traversent les tissus et atteignent les ovules. Les œufs, enfermés dans un cocon, ont un développement direct, sans trochophore.
Régénération et reproduction asexuée
Les Polychètes et les Oligochètes manifestent un pouvoir de régénération développé, alors que les Hirudinées en sont totalement dépourvues. Il est faux, cependant, de croire que, coupé en deux, un Ver de terre donne deux individus viables ; les recherches expérimentales entreprises ont montré que les choses n’étaient pas si simples !
Plus la section transversale d’un Lombric est postérieure, moins la régénération de la tête est facile ; de même, la région postérieure ne se reforme que si la section n’est pas trop antérieure ; il y a une zone — située entre les segments 15 et 30 — où aucune régénération n’est possible ; donc, à partir d’un Ver coupé, on obtient, au mieux, la néo-formation d’une seule extrémité. Par contre, chez Lumbriculus, il y a une zone moyenne où coexistent les deux potentialités de régénération ; isolée, cette zone produit un individu complet. Chez Eisenia fœtida, il faut que les segments 9 à 20 soient présents pour obtenir un Ver viable.
Des cellules indifférenciées, les néoblastes, d’origine mésodermique, se trouvent sur les dissépiments ; elles migrent vers le lieu de l’amputation, se divisent en donnant un blastème de régénération, à partir duquel s’édifient de nouveaux éléments mésodermiques (cœlome, muscles, néphridies) ; l’ectoderme proche de la section fournit le tégument et les ganglions nerveux ; l’intestin prolifère dans le régénérât. Le système nerveux exerce une influence certaine dans les processus de régénération.
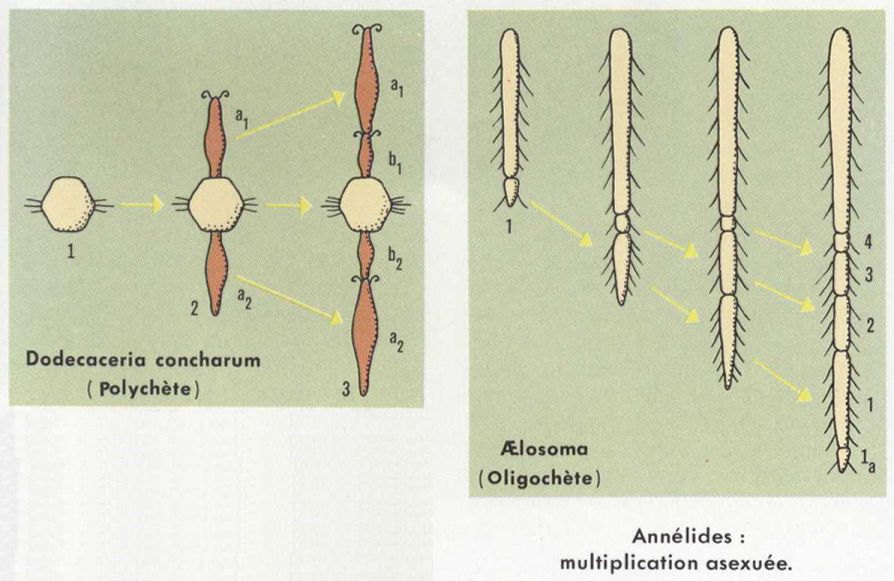
La reproduction asexuée est répandue dans les groupes où la régénération est active ; elle se manifeste selon des modalités très variées, que l’on assimile soit à la scissiparité, soit au bourgeonnement.
Le corps du Ver peut se fragmenter, et chaque tronçon donne un individu complet ; à la limite, l’animal égrène ses métamères un par un (Lumbriculus, Oligochète ; Dodecaceria, Polychète). Le Ver peut se subdiviser en plusieurs zones, qui restent pendant un temps en file linéaire avant de se séparer et de régénérer les parties manquantes (Æolosoma, Oligochète ; Ctenodrilus, Polychète). Dans d’autres cas, la régénération a heu avant la séparation des stolons (Dero, Oligochète ; Salmacina, Polychète). Enfin, le bourgeonnement peut intervenir en relation avec la dissémination des gamètes, les stolons se chargeant seuls des produits génitaux : le Polychète Autolytus effectue un bourgeonnement en chaîne ; la souche est asexuée, et les stolons présentent un dimorphisme sexuel accentué.