vision (suite)
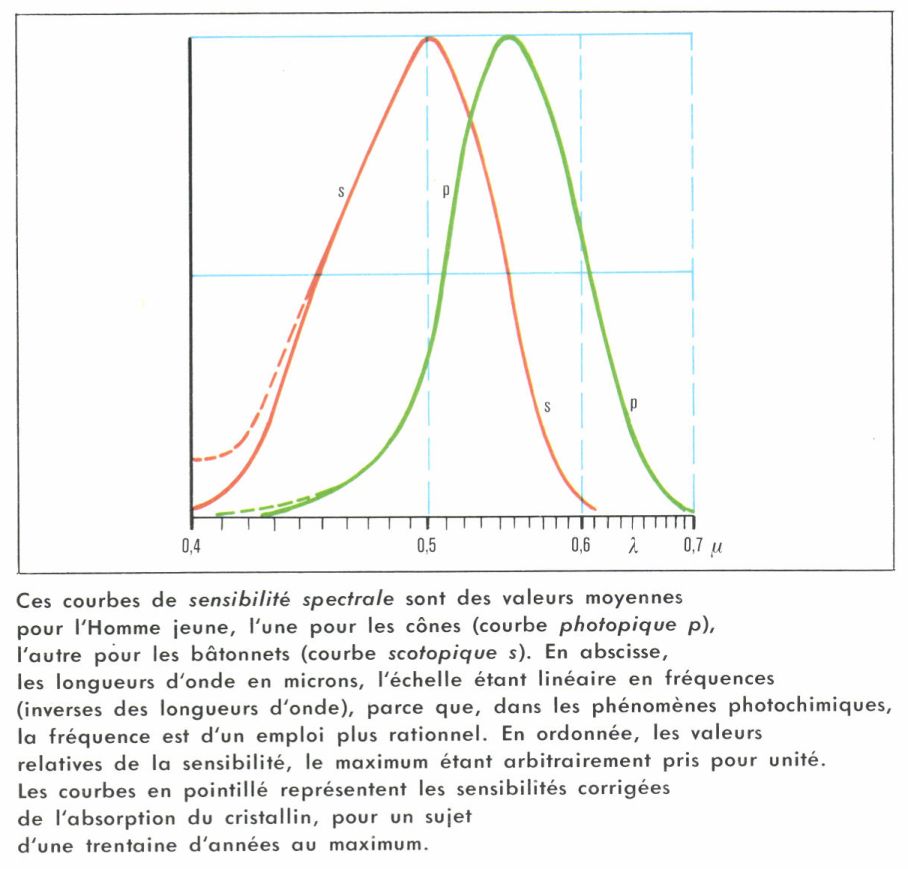
La longueur d’onde λm du maximum de la courbe V (λ) est en moyenne à 507 nm (le nanomètre [nm], autrefois nommé millimicron, est égal au millionième du millimètre) pour la courbe scotopique des bâtonnets et à 555 nm pour la courbe photopique des cônes. Les courbes en cloche descendent régulièrement de part et d’autre et s’annulent pratiquement (V = 0,001) à 385 et à 645 nm, d’une part, et à 408 et à 721 nm, de l’autre ; en chiffres ronds, les limites du visible sont de l’ordre de 400 et de 700 nm, ce qui est assez étroit : moins d’une octave pour parler comme les musiciens.
Ces résultats sont relatifs à l’Homme, mais il semble que, pour tous les Vertébrés, il n’y ait, en gros, qu’une seule lumière ; nous reviendrons plus loin sur les Insectes. On a bien entendu longuement disserté sur la raison biologique de la position du visible dans l’énorme domaine des radiations électromagnétiques : il y a une évidente relation avec le rayonnement solaire, tel qu’il nous arrive, tamisé par l’atmosphère et éventuellement par la mer selon l’habitat de l’animal.
À vrai dire, la rétine peut répondre à des radiations extérieures au domaine visible : ainsi, l’ultraviolet, de 365 nm par exemple, est vu par un sujet jeune avec une couleur bleuâtre, probablement par fluorescence des milieux de l’œil ; avec les années, le cristallin jaunit et absorbe de plus en plus les courtes longueurs d’onde. La sensibilité à l’ultraviolet disparaît ; elle revient fortement chez les sujets aphakes, à qui on a enlevé le cristallin à la suite d’une cataracte. Les rayons X sont, eux aussi, faiblement perçus, et il semble même que les rayons cosmiques soient responsables des éclairs décrits par les astronautes.
Pigments visuels
On a fait d’innombrables hypothèses sur le mécanisme de la vision ; actuellement, on sait que c’est un phénomène photochimique : dans les bâtonnets et les cônes, les quanta de lumière (énergie élémentaire du rayonnement) sont absorbés par une substance dite pigment, et les modifications induites dans le pigment sont à l’origine du message nerveux. Il faut, évidemment, que cette modification soit réversible pour que la vision continue.
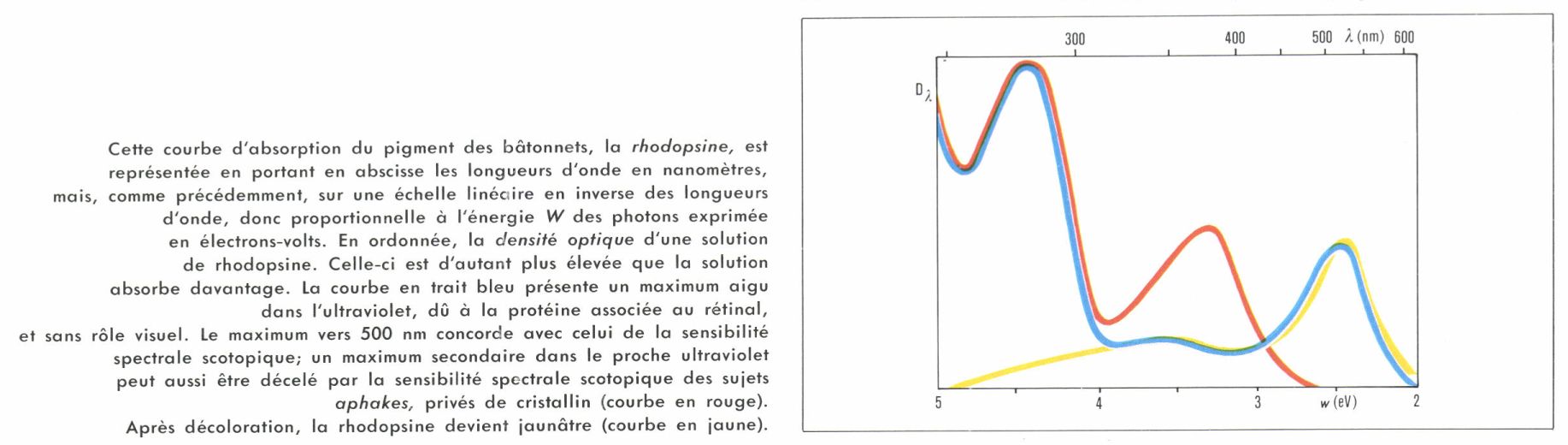
La question est résolue pour les bâtonnets. En décollant une rétine de Grenouille, Franz Christian Boll constata, en 1876, que sa couleur rose jaunissait à la lumière ; elle contient donc une substance photosensible, qu’il appela pourpre rétinien et qu’on nomme maintenant rhodopsine. Dès l’année suivante, on montra l’existence du pourpre dans la rétine d’un criminel exécuté dans l’obscurité. En 1903, Konig prouva que la courbe d’absorption du pourpre humain est voisine de la courbe d’efficacité lumineuse scotopique, ce qui démontre le rôle de ce pigment dans la vision par les bâtonnets.
La rhodopsine a provoqué d’innombrables travaux, qui culminent avec l’œuvre de George Wald (né en 1906), professeur à Harvard, prix Nobel en 1967. Elle est constituée par l’union d’une protéine spéciale, dite opsine, avec un chromophore, appelé rétinal, responsable de l’absorption dans le visible et qui est l’aldéhyde d’un alcool non saturé, la vitamine A, ou rétinol. On savait depuis longtemps que la carence en vitamine A dans l’alimentation provoquait des troubles dans la vision de nuit. Wald a montré que l’action photochimique de la lumière est simplement une photo-isomérisation du rétinal : pour être sensible à la lumière, le rétinal des bâtonnets doit exister non pas sous la forme stable trans, où tous les atomes de carbone d’une chaîne à simples et à doubles valences alternées sont alignés, mais sous une certaine forme cis, où la chaîne est tordue après un carbone bien défini. L’absorption d’un photon* (quantum de lumière) déclenche le retour de la forme cis à la forme trans ; c’est le seul effet de la lumière, tout le reste étant une suite compliquée de transformations chimiques qui modifient l’équilibre électrique dans le bâtonnet ; d’où un potentiel très faible qui sera l’origine de l’influx nerveux. (Il est intéressant de rappeler que ce rôle des structures des isomères chimiques dans les phénomènes de la vie a débuté par les travaux de Pasteur.) Le retour du rétinal à la forme cis se produit sous l’effet d’une enzyme sécrétée par l’épithélium pigmentaire qui tapisse l’arrière de la rétine contre les bâtonnets.
Puisque la probabilité qu’un photon de lumière soit absorbé est proportionnelle à l’ordonnée de la courbe d’absorption de la rhodopsine, on comprend que celle-ci doive s’identifier avec la courbe d’efficacité lumineuse scotopique. En réalité, il faut, d’une part, tenir compte des écrans colorés (le cristallin en particulier) interposés avant la rétine et, d’autre part, évaluer la courbe d’efficacité lumineuse non pas en énergies, mais en nombre de photons, en se rappelant que l’énergie est inversement proportionnelle à la longueur d’onde.
Le spectre d’absorption de la rhodopsine peut se déterminer au laboratoire ; pour cela, on met la rhodopsine d’une rétine détachée de l’œil en solution dans de la digitonine. On peut, d’une façon plus directe mais plus difficile, faire la mesure in situ : en 1897, Abelsdorf avait constaté, chez des sujets albinos, qui manquent d’épithélium pigmentaire absorbant derrière leur rétine, ou chez des animaux à tapis réfléchissant (v. œil), qu’on pouvait à l’ophtalmoscope voir la coloration rose de la rétine riche en pourpre et en suivre la décoloration à la lumière. Depuis 1951, cette méthode a permis, même chez des sujets normaux, de suivre la variation de transparence de la rétine pendant la décoloration de la rhodopsine. Ces deux techniques concordent, et l’on retrouve bien la courbe d’efficacité scotopique.
Pour le cas des cônes, le problème est beaucoup moins avancé, et l’on n’a pas pu, jusqu’ici, isoler de pigments avec certitude ; nous reviendrons là-dessus à propos de la vision des couleurs. Cependant, dans la fovéa, où il n’y a pas de bâtonnets, la méthode ophtalmoscopique conduit à une courbe spectrale d’absorption qui, en gros, rappelle la courbe d’efficacité photopique, ce qui est un argument en faveur de la théorie photochimique de la vision par les cônes, comme pour les bâtonnets, mais avec des pigments différents, ce qui explique l’effet Purkinje. Une donnée intéressante est la mesure comparée des absorptions maximales après complète adaptation à l’obscurité, c’est-à-dire quand les pigments se sont accumulés tant dans les bâtonnets que dans les cônes. Vers 500 nm dans le premier cas et 560 nm dans l’autre, ces absorptions sont du même ordre, à savoir 0,2, ce qui signifie que deux photons sur dix sont absorbés en moyenne. Cela prouve que la différence de sensibilité (le rapport des seuils est en gros de 1 à 1 000) entre bâtonnets et cônes ne provient pas d’une absorption plus forte par les premiers ; c’est une question de sommation : les cônes de la fovéa sont presque indépendants ; au contraire, de nombreux bâtonnets se groupent en une seule unité réceptrice sur une même cellule ganglionnaire par l’intermédiaire des bipolaires (v. œil) : il suffit qu’un petit nombre de photons, peut-être deux seulement, tombent en même temps (à 0,01 s près) dans la même unité réceptrice pour qu’un message parte vers le cerveau, et le rapport des seuils mesure celui des surfaces des unités réceptrices et non celui des sensibilités individuelles des récepteurs. Nous reviendrons sur ce point à propos de l’acuité visuelle.