




Angiospermes (suite)
Les Angiospermes sont en majorité des espèces ayant des racines, des tiges, des feuilles et des fleurs. Ce ne sont que les adaptations secondaires à des milieux extrêmes (désertique ou aquatique) ou à une biologie très particulière (parasitisme) qui provoquent l’atrophie, voire la suppression, de certains organes : réduction du système foliaire pour les plantes désertiques, suppression de la racine pour les parasites.
La morphologie et la structure des racines, des tiges et des feuilles caractérisent les Angiospermes d’une manière moins rigoureuse que les fleurs. Alors que les Gymnospermes ont surtout des aiguilles persistantes (il y a bien des exceptions, tel le Ginkgo à feuilles aplaties et caduques), chez les Angiospermes, les feuilles ont ordinairement la forme de lames vertes plus ou moins découpées, qui ne vivent le plus souvent que l’espace d’une saison (mais les feuilles de Romarin, par exemple, sont très étroites et persistent plusieurs années) [v. aussi feuille, racine, tige].
Reproduction sexuée
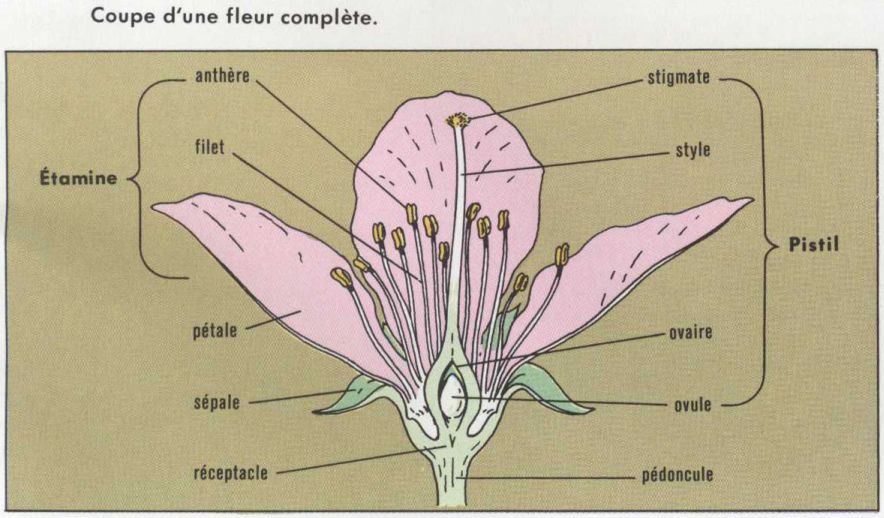
Les organes reproducteurs ont chez les Angiospermes l’évolution la plus poussée et ce sont eux, par leur beauté, leur structure, et surtout leur biologie, qui caractérisent ce sous-embranchement. Mais on rencontre de très importantes variations dans la morphologie et même dans la structure de la fleur*.
La fécondation
Le transport du grain de pollen de l’étamine sur le stigmate, puis la pénétration du tube pollinique (germination du grain de pollen) dans les tissus femelles jusqu’à l’oosphère sont chez les Angiospermes les premiers actes de la fécondation, dont l’essentiel est la fusion entre les gamètes mâles et femelles.
Une fois le tube pollinique arrivé au niveau des synergides du sac embryonnaire, le noyau végétatif se résorbe, et il ne reste plus alors que les deux gamètes mâles ordinairement nus, sans cils, qui se déversent dans le sac, l’extrémité du tube pollinique étant alors gélifiée.
À ce moment, les deux noyaux polaires du sac embryonnaire vont ordinairement fusionner pour former un noyau à 2 n chromosomes (noyau secondaire), qui alors s’unit à un des deux anthérozoïdes pour donner l’œuf accessoire à 3 n chromosomes, lui-même origine de l’albumen*.
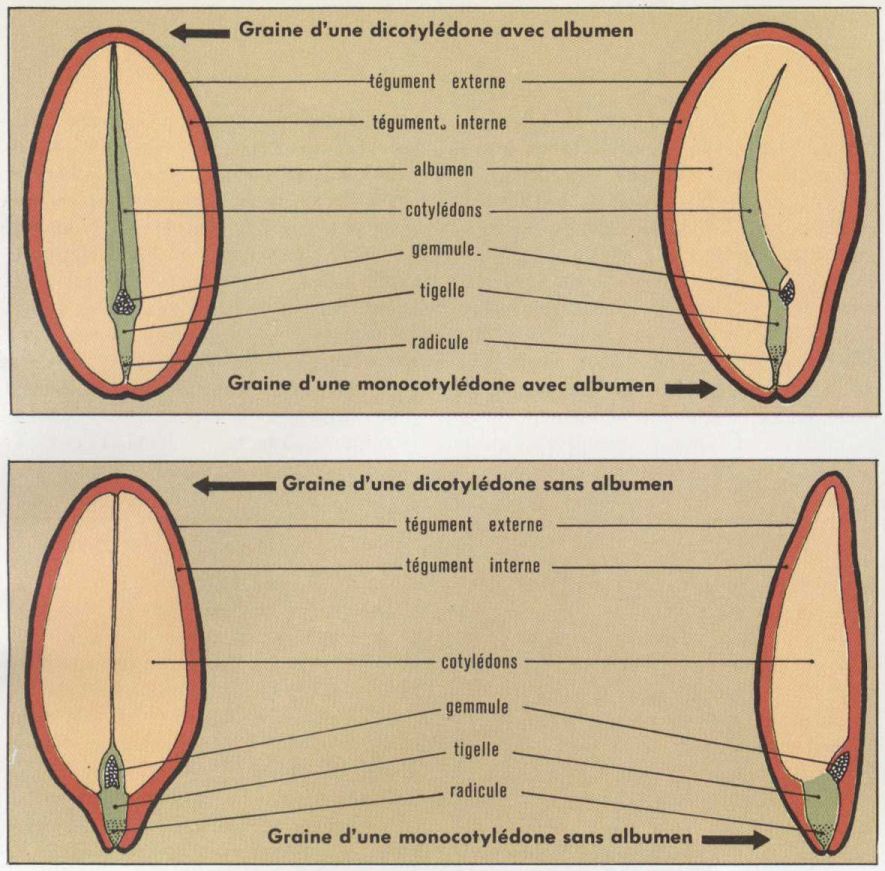
Enfin, le dernier noyau du sac embryonnaire, très riche en substance chromosomique et le plus souvent disposé au-dessous des synergides, va fusionner avec le deuxième anthérozoïde pour donner l’œuf vrai, qui après de nombreux cloisonnements fournira l’embryon de la nouvelle plante. Cette fécondation est très sensiblement la même dans tout le sous-embranchement des Angiospermes (sauf de très rares exceptions : Fagacées), et elle s’oppose ainsi à celle des Gymnospermes, qui n’ont pas d’albumen. Une fois ces phénomènes de fécondation terminés, l’ovule puis l’ovaire subissent d’importantes transformations, qui les font respectivement devenir graine* et fruit. Le complet développement de la graine s’effectue à l’intérieur même de l’ovaire : c’est une des caractéristiques des Angiospermes.

Origine des Angiospermes
Ces plantes, qui sont probablement les plus récentes des Phanérogames, ont leurs représentants les plus anciennement connus dans le Jurassique supérieur. Cependant, dès le Crétacé, de nombreux restes de végétaux Dicotylédones ont pu être identifiés et rapportés à des familles actuelles : Araliacées, Euphorbiacées, Lauracées, Légumineuses, Magnoliacées, Myrtacées ; Moracées, Fagacées ; Ébénales, Éricales et Primulales. Enfin, comme Monocotylédones, on a remarqué des espèces des familles des Aracées, Cypéracées, Liliiflores et Palmiers. L’origine de ces espèces doit être recherchée bien plus dans les groupes des Gymnospermes primitives (Cycadales et Ginkgoales) que chez les Cryptogames vasculaires. En effet, certaines Angiospermes possèdent des particularités anatomiques voisines de celles des Gymnospermes primitives. D’autres caractéristiques archaïques quant à l’organisation florale (Pipéracées, Résédacées, Violariacées) militent aussi en faveur de cette thèse. Enfin, on pense que l’origine des Angiospermes ne serait pas monophylétique, mais polyphylétique : les Angiospermes ne dériveraient pas toutes des mêmes Gymnospermes, mais les unes des Bennettitales, les autres des Cordaïtales, l’ancêtre commun étant encore plus primitif (Cryptogames vasculaires archaïques).
J.-M. T. et F. T.