




cellule (suite)
Les centrioles
Il existe, chez les Métazoaires principalement, un couple de centrioles au voisinage du noyau. Ils se présentent comme de petits cylindres, orthogonaux entre eux, dont les génératrices sont faites de microtubules disposés en neuf groupes de trois (triplets). Ces centrioles sont susceptibles de polariser dans leur direction des protéines fibreuses de l’hyaloplasme, notamment lors de la division cellulaire, où se réalise un important fuseau de microtubules paraissant intervenir dans la répartition du matériel nucléaire entre les deux cellules filles. Sous le plasmalemme peuvent exister des organites tout à fait analogues aux centrioles, les cinétosomes, qui sont chacun à l’origine d’un flagelle, ou cil. Deux des microtubules de chaque triplet s’allongent ; il se constitue ainsi un cylindre de neuf doublets qui soulèvent en doigt de gant la membrane plasmatique. Ce système est renforcé d’un couple de microtubules axiaux. Les flagelles sont mobiles ; ils peuvent onduler diversement, mais on ignore la nature des processus responsables de leurs mouvements.
Le noyau
Très généralement, la cellule eucaryote ne comporte qu’un seul noyau, souvent sphérique.
La membrane nucléaire ne représente qu’une différenciation locale du réticulum endoplasmique général, avec lequel elle est, par places, en continuité. Elle est faite de deux membranes unitaires parallèles qui ménagent à intervalles réguliers des perforations, ou pores nucléaires, assurant le transit de molécules relativement grosses de part et d’autre.
Le suc nucléaire, ou nucléoplasme, renferme des enzymes (nucléases et estérases notamment) et de l’A. R. N. partiellement sous forme particulaire ou filamenteuse.
Les chromosomes sont faits de très longues molécules d’acide désoxyribonucléique (A. D. N.), auxquelles sont associées des protéines (histones notamment). Dans l’intervalle des périodes de division cellulaire (interphase), ces fibres nucléoprotéiques sont étirées, entremêlées par places (blocs chromatiques). C’est seulement lors de la division cellulaire qu’elles se raccourcissent en se spiralisant sur elles-mêmes. Les chromosomes se présentent alors comme des bâtonnets et montrent nettement leur individualité : leur nombre, leur forme et leurs dimensions respectives sont caractéristiques de l’espèce. Ils représentent en effet le support essentiel des caractères héréditaires de la cellule et déterminent, par l’intermédiaire d’acides ribonucléiques synthétisés à leur contact, la constitution spécifique des protéines de structure. Le ou les nucléoles représentent précisément des sites où s’accumulent les A. R. N. d’origine chromosomienne. Ils dépendent d’un segment chromosomique (organisateur nucléolaire) qui les traverse ou auquel ils sont accolés. Ils sont faits d’une matrice dense, non limitée par une membrane et au sein de laquelle s’enchevêtrent des fibrilles et s’accumulent des granules de 150 Å. La matrice est protéique, les granules ont la même constitution que les ribosomes et les fibrilles représentent des molécules d’A. R. N. Une partie de ce matériel est destinée à pourvoir le système ribosomien du cytoplasme, tandis qu’une autre reste sur place et assure les synthèses protidiques du noyau. Chez les Protocaryotes, le système nucléaire est limité à un unique chromosome baignant en plein cytoplasme et fait d’une seule molécule d’A. D. N. circulaire.
Biosynthèse des protéines
Ce sont les molécules d’A. D. N., essentiellement celles des chromosomes et en partie celles des mitochondries et des plastes, qui sont responsables du maintien de la constitution spécifique des protéines cellulaires. Elles servent en quelque sorte de moule pour la synthèse des différents types d’A. R. N., en présence d’enzymes (A. R. N. polymérase). Parmi ces molécules d’A. R. N., les unes, très hautement polymérisées (poids moléculaire : 3 × 106 environ), constituent, après combinaison avec des protéines, les ribosomes du cytoplasme. D’autres, beaucoup plus petites (A. R. N. soluble ou de transfert) [poids moléculaire : 30 000 environ], diffusent dans le cytoplasme et sont susceptibles de servir chacune de support à un acide aminé déterminé, qu’elles déposent à la surface des ribosomes. Enfin, le long des molécules d’A. D. N., certaines régions, qualifiées de gènes et présentant une séquence déterminée de bases puriques et pyrimidiques, assurent le codage de molécules d’A. R. N. d’un poids moléculaire intermédiaire de l’ordre de 500 000. Cet A. R. N., après transit dans le nucléole, migre (A. R. N. messager) au travers des pores de la membrane nucléaire, et c’est sans doute lui que l’on peut repérer dans l’hyaloplasme sous forme de petits filaments hélicoïdaux sur lesquels se fixent des ribosomes. Grâce à l’énergie fournie par l’A. T. P. cytoplasmique, les acides aminés, portés par le ribosome, se combinent en une chaîne polypeptidique selon des séquences déterminées par celles des bases de l’A. R. N. messager voisin. Plusieurs ribosomes pouvant se fixer le long d’une même molécule d’A. R. N. messager, les polypeptides synthétisés par chacun d’entre eux s’unissent, selon un ordre défini, pour constituer de grosses molécules protidiques spécifiques.
Division cellulaire
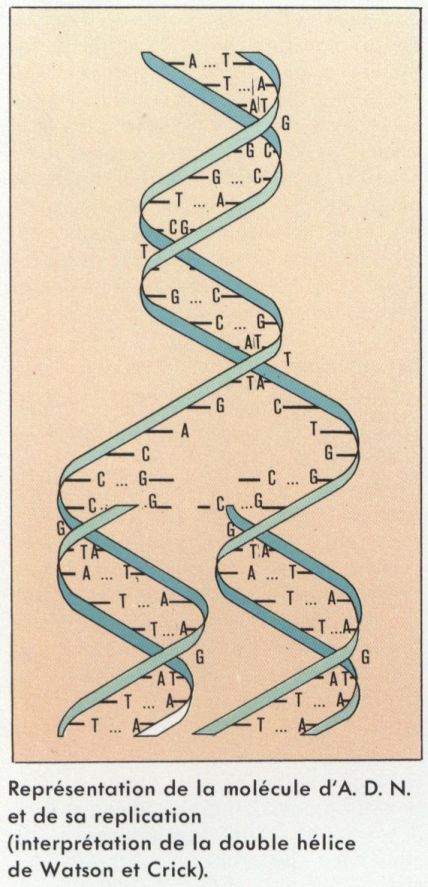
La reproduction, caractère fondamental de tout être vivant, et la croissance des organismes pluricellulaires résultent essentiellement de l’aptitude de leurs cellules à se diviser, c’est-à-dire à donner naissance à de nouvelles cellules identiques à elles-mêmes. En dernière analyse, ce processus de multiplication cellulaire a pour origine la duplication des A. D. N. de leurs noyaux, ces derniers conditionnant par la suite, par les synthèses qu’ils déterminent, la duplication de l’ensemble des constituants de la cellule. Chaque molécule d’A. D. N. est faite de deux chaînes polynucléotidiques, torsadées l’une autour de l’autre en une double hélice et appariées de façon complémentaire au niveau de chacun de leurs radicaux basiques, la thymine s’associant à l’adénine, la cytosine à la guanine. La duplication de cet ensemble a lieu dans l’intervalle de deux divisions cellulaires, ou interphase ; elle résulte de la séparation des deux brins, chacun d’eux induisant à son contact la synthèse d’un brin qui lui est complémentaire et auquel il s’associe. L’autoduplication de l’A. D. N. de chaque chromosome entraîne celle du chromosome lui-même en deux éléments fils, ou chromatides, encore associés. Par la suite, la division nucléaire, ou mitose, assurera la répartition équationnelle de ces chromatides entre les deux noyaux fils : dans un premier temps de la mitose, ou prophase, les couples de chromatides se raccourcissent en se spiralisant, tandis que, généralement dans le cytoplasme, un nouveau couple de centrioles apparaît à proximité du premier et s’en écarte pour occuper par rapport au noyau une position diamétralement opposée. À la fin de la prophase, la membrane nucléaire se fragmente généralement : des faisceaux de microtubules, ou fuseaux, se tendent entre les deux couples centriolaires, et les chromatides associées se fixent à ces microtubules par une région spécialisée (centromère). À l’étape suivante (métaphase), tous les centromères se disposent à l’équateur du fuseau ; le raccourcissement des microtubules entraîne la séparation des chromatides jumelles (anaphase) et leur ascension en direction des pôles fusoriaux. Se réalise ainsi une répartition équationnelle en deux lots de chromatides qui s’entremêlent et s’étirent en se déspiralisant (télophase). Des vésicules ergastoplasmiques entourent chacun des noyaux fils en cours de reconstitution et, en devenant coalescentes, elles reforment de nouvelles membranes nucléaires. Finalement a lieu la division cellulaire proprement dite, ou cytocinèse, par étranglement équatorial de la cellule mère.