Jamais infidèle à lui-même, à ses obsessions et à son goût de l’esthétisme grandiose, Luchino Visconti est un des cinéastes qui a le mieux parlé d’un passé politique à résonances contemporaines, et sa célébration de la beauté ne l’a jamais empêché d’être lucide et virulent. Il peut même être considéré comme un des plus grands, puisqu’il n’a jamais, en fin de compte, parlé que du temps, de sa fuite et de la mort, que même l’œuvre d’art ne peut arrêter dans sa course.
M. G.
G. Ferrara, Luchino Visconti (Seghers, 1964 ; nouv. éd., 1970). / Y. Guillaume, Visconti (Éd. universitaires, 1967).
vision
Fonction qui procure aux animaux une information sur les objets par l’intermédiaire de radiations électromagnétiques situées dans le domaine de longueurs d’onde dit lumière*.
La psychologie classique distinguait dans la vision les sensations* (lumière, couleur) et les perceptions* (forme, détails, mouvements, relief, etc.). Cette division, reflet du dualisme corps-esprit, s’est atténuée : pour le physiologiste actuel, la vision met en jeu une chaîne nerveuse très élaborée qui part des cellules photosensibles de la rétine (v. œil) pour aboutir, après divers relais, au cortex visuel (v. cerveau) ; de là, le message nerveux, après des parcours complexes dans plusieurs régions cérébrales, revient vers la périphérie, en particulier pour commander certains réglages comme l’ouverture de la pupille de l’œil, qui agit sur la lumière admise, et l’accommodation, qui, en modifiant la forme du cristallin, permet la mise au point précise de l’image sur la rétine. Évidemment, certaines perceptions, celle du relief par exemple, ont, comme nous le verrons, une origine centrale, dans le cerveau, mais d’autres, telles celles des formes et des mouvements, mettent déjà en œuvre des mécanismes rétiniens, donc périphériques, ce qui rend illusoire l’ancienne classification de la psychologie.
Plus le système nerveux se perfectionne dans la série animale et plus les fonctions visuelles s’enrichissent. Aussi est-ce chez les Mammifères, et surtout chez l’Homme, que nous étudierons la vision, mais nous dirons quelques mots, pour terminer, de la vision des Insectes, qui pose des problèmes remarquables et encore mal élucidés.
Sensibilité lumineuse
Lorsqu’une surface S de la rétine d’un animal reçoit pendant un temps t une radiation électromagnétique de longueur d’onde λ bien définie et transportant à travers cette surface une énergie totale W, l’animal peut ou non réagir. On appelle seuil* le cas limite, donc la plus faible lumière produisant une réponse. Sur l’animal, on peut déterminer le seuil par conditionnement : on dresse l’animal à répondre (en appuyant par exemple sur une pédale quand il voit la lumière) et on le récompense par de la nourriture quand il répond à juste titre, tandis qu’on le punit par une décharge électrique s’il répond à tort et à travers. Une autre technique repose sur l’électrophysiologie : nous y reviendrons plus loin. Chez l’Homme adulte, la réponse est donnée par le sujet lui-même, qui dit s’il voit ou non quelque chose, et on élimine les réponses douteuses par un traitement statistique approprié.
Les lois du seuil lumineux sont complexes : si t est très petit, inférieur à un centième de seconde chez l’Homme, W mesure le seuil : on dit qu’il y a sommation temporelle pour les éclairs brefs ; pour des expositions dépassant plusieurs secondes, c’est au contraire le quotient W/t qui intervient, donc la puissance énergétique (reçue par la surface S), laquelle se mesure en watts. De même, si S est petit — si, par exemple, la source est vue sous un diamètre apparent moindre que l’ d’angle (source ponctuelle) —, il y a sommation spatiale, et W intervient seul, tandis que, pour de grandes surfaces, le quotient W/S est la variable intéressante.
Chez beaucoup d’animaux, et chez l’Homme en particulier, la rétine contient deux types de cellules sensibles à la lumière, les cônes et les bâtonnets (v. œil), ces derniers étant absents dans une région centrale de la rétine dite fovéa. Cette dualité anatomique s’accompagne d’une dualité fonctionnelle : les cônes, qui servent à la vision diurne (photopique), sont moins sensibles que les bâtonnets, qui servent à la vision nocturne. En outre, seuls les cônes assurent la vision des couleurs, si l’animal la possède.
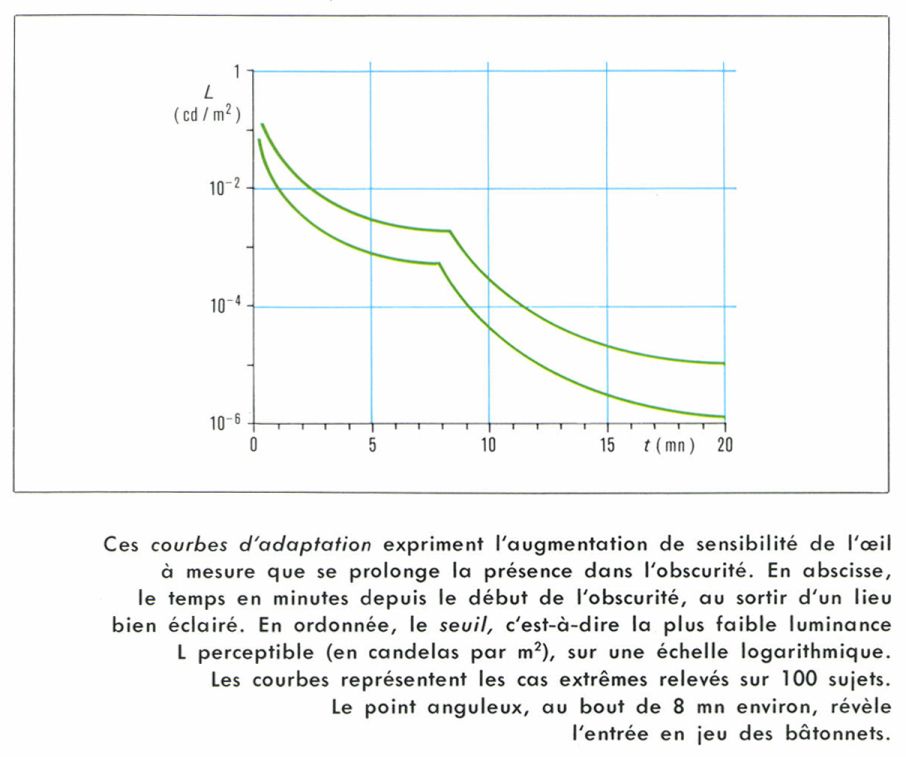
Du fait de la dualité rétinienne, les effets d’adaptation compliquent la question de mesure des seuils : quand on pénètre dans une cave, on ne voit d’abord rien, puis l’œil s’accoutume à la faible lumière, et, au bout de vingt minutes environ, on est bien adapté ; le seuil ne change alors plus guère.
Les courbes d’adaptation (seuil en fonction du temps de présence à l’obscurité) présentent au bout de quelques minutes un point anguleux qui révèle le passage de la vision des cônes à celle des bâtonnets, qui s’adaptent mieux et deviennent de ce fait plus sensibles.
Sensibilité spectrale
Si l’on détermine le seuil en fonction de la longueur d’onde λ de la radiation qui excite la rétine, on constate que l’énergie nécessaire W varie beaucoup : elle est minimale au milieu du spectre visible et devient considérable quand on approche de l’ultraviolet et de l’infrarouge, qui continuent le visible vers les courtes et grandes longueurs d’onde. On définit la sensibilité spectrale, ou efficacité lumineuse, par une grandeur V (λ) inversement proportionnelle à W. On convient de faire V = 1 pour la longueur d’onde à laquelle l’œil est le plus sensible ; l’efficacité est donc toujours inférieure à 1.
La sensibilité spectrale n’est pas la même pour les cônes et les bâtonnets, ces derniers ayant un maximum décalé vers les courtes longueurs d’onde. On peut le constater aisément en cherchant par exemple un papier rouge et un papier bleu qui, en bonne lumière, semblent aussi lumineux l’un que l’autre ; si le jour tombe, l’œil s’adapte à l’obscurité ; bientôt les bâtonnets supplantent les cônes, les couleurs disparaissent, le papier bleu devient gris clair, et le rouge presque noir. C’est ce qu’on appelle l’effet Purkinje en l’honneur du physiologiste tchèque Jan Evangelista Purkynĕ (1787-1869). Pour les bâtonnets, la détermination de V (λ) est aisée : le sujet, bien adapté à l’obscurité, regarde un champ photométrique comprenant deux plages adjacentes, par exemple deux demi-cercles séparés par un diamètre aussi fin que possible ; une plage reçoit une lumière de comparaison fixe, qui peut être de la lumière blanche, comprenant toutes les radiations visibles ; l’autre reçoit une lumière de longueur d’onde λ variable et d’énergie réglable. On demande au sujet d’égaliser la luminosité des plages. Pour éviter l’entrée en jeu des cônes, il faut utiliser des lumières faibles (le mieux serait le seuil lui-même, mais alors la comparaison est imprécise) et employer la région de la rétine la plus riche en bâtonnets, vers 10° du point de fixation qui est au centre de la fovéa. Pour les cônes, il y a des difficultés provenant de la gêne du sujet à comparer les luminosités de plages de couleurs différentes ; on les surmonte par des techniques (méthodes pas à pas ou papillotement) que le lecteur trouvera dans les ouvrages de photométrie.