




toucher ou tact (suite)
• Nocicepteurs thermiques. En réalité, comme les nocicepteurs mécaniques, ils sont excités par de fortes stimulations mécaniques, mais ils répondent de plus à une augmentation importante de température de la peau (et moins souvent à une diminution). Les fibres identifiées sont souvent de type C avec une vitesse inférieure à 2,5 m/s ; la figure 13 donne un exemple des réponses d’une telle fibre. Mais, quoique moins nombreuses, des fibres myélinisées dont la vitesse est supérieure à 7 m/s interviennent également.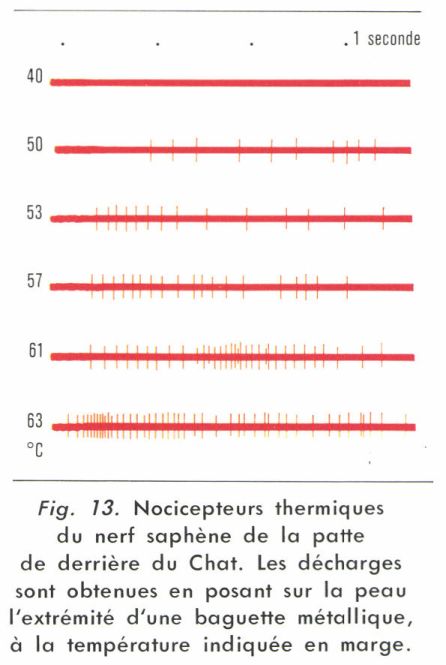
Qu’il s’agisse des nocicepteurs mécaniques ou mécanothermiques, on n’a pas identifié avec certitude les terminaisons, dont on pense qu’elles pourraient être situées pour la plupart très superficiellement, contre la couche basale de l’épiderme, et peut-être à l’intérieur même de cette couche.
• La double douleur. On aura remarqué que, pour les deux types de nociception, on trouve un contingent de fibres myélinisées dont la vitesse de conduction varie de 5 à 50 m/s et un contingent de fibres sans myéline dont la vitesse de conduction est de l’ordre de 1 m/s. Ce fait est à rapprocher du phénomène que l’on peut observer, par exemple, lorsqu’on se pince accidentellement ou lorsqu’on plonge la main dans une eau trop chaude (65 °C) : à une première douleur aiguë, brève et bien localisée, qui provoque le retrait rapide de la main, succède, avec un retard de l’ordre de 1 seconde, un « élancement » diffus, une douleur plus sourde, plus difficile à supporter. Si l’on admet que la stimulation douloureuse active simultanément les terminaisons des fibres relativement rapides et celles des fibres lentes, on conçoit que le message qui aura emprunté ces dernières parvienne aux centres nerveux avec retard. En prenant une vitesse moyenne de 25 m/s pour les fibres rapides, on peut calculer que, sur une distance de 1 m, le message prend un retard de 0,96 seconde. Cette explication de la double douleur, si elle n’est pas unanimement admise, recueille cependant la majorité des suffrages.
La genèse du message sensoriel
Pour qu’une stimulation soit efficace, il faut qu’elle provoque la naissance d’un potentiel de récepteur (v. sensation). Nous étudierons cette opération, dite de transduction, en prenant comme exemple le corpuscule de Pacini, dont la structure et le fonctionnement ont fait l’objet de nombreux travaux. À partir du potentiel de récepteur s’élabore ensuite le message sensoriel, constitué de potentiels d’action, message codé dont l’information dépend de la nature du récepteur ; nous le démontrerons en comparant les codages dont sont capables le corpuscule de Pacini et les complexes de Merkel du dôme de Iggo. Nous envisagerons ensuite brièvement, car les informations sont rares, le mécanisme de la transduction dans les thermorécepteurs et les nocicepteurs.
La transduction dans un mécanorécepteur, le corpuscule de Pacini
La figure 14 donne le schéma d’ensemble et le détail de la massue terminale du corpuscule. On notera que la fibre ne perd sa myéline qu’après être entrée dans l’enveloppe, en sorte qu’un nœud de Ranvier se trouve inclus dans le corpuscule. L’enveloppe conjonctive enferme deux capsules concentriques : la capsule externe, formée de feuillets continus emprisonnant entre eux un fluide visqueux, et la capsule interne, formée d’hémilamelles, représentées sur le schéma reconstitué d’après les observations au microscope électronique.
Les physiologistes ont expérimenté sur des corpuscules de Pacini isolés, avec un fragment de leur fibre nerveuse, à partir du péritoine du Chat, où ceux-ci apparaissent à l’œil nu comme des grains de mil dans la transparence des feuillets mésentériques.
En réponse à un choc mécanique de quelques microns d’amplitude appliqué sur l’enveloppe, le corpuscule fournit une double réponse électrique, reproduite sur la figure 15 A. Si l’on ajoute de la procaïne au sérum physiologique dans lequel baigne la préparation, les corpuscules rapides de la double réponse sont éliminés (fig. 15 B) : il s’agit de potentiels d’action dont on sait qu’ils sont bloqués par la procaïne (d’où le rôle anesthésique de ce produit). La variation électrique qui subsiste constitue le potentiel de récepteur, premier élément électrophysiologique provoqué par la stimulation.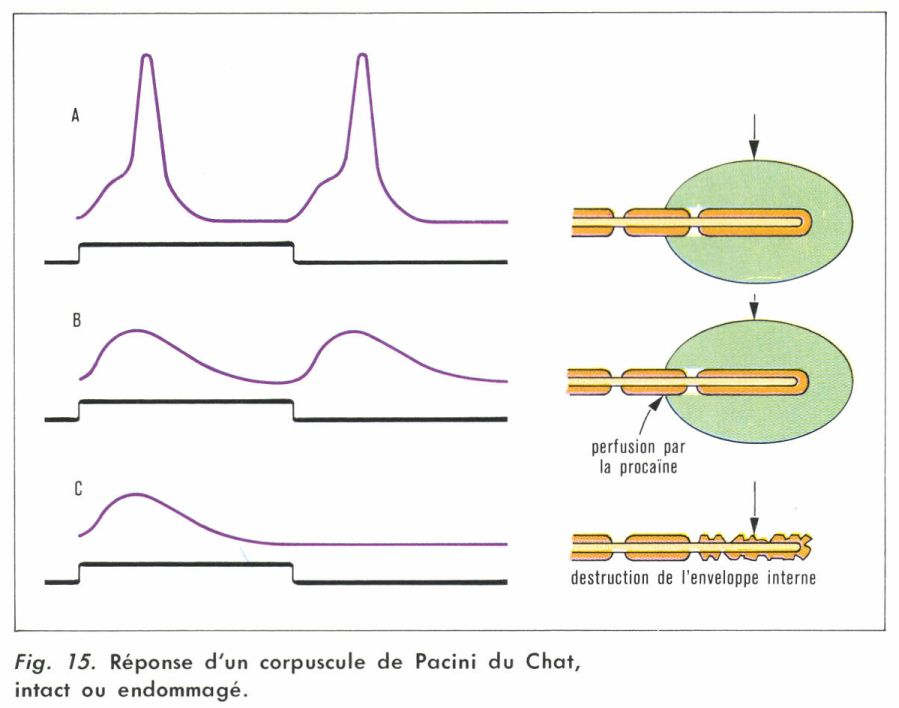
Si l’on débarrasse la fibre nerveuse de la capsule externe — et, ce faisant, on endommage également la capsule interne —, le même choc, appliqué directement (fig. 15 C), ne provoque plus de réponse à la cessation de la déformation. Celle-ci était due en effet à l’action de l’enveloppe et de la capsule externe élastique — qui, libérées, reprenaient leur forme initiale en exerçant ainsi sur la fibre une pression orthogonale à la pression subie initialement.
Quant au potentiel de récepteur, on a pu montrer qu’il était le résultat d’une entrée d’ions sodium (Na+) dans la fibre, entrée rendue possible par la déformation mécanique de la membrane qui entraîne sa perméabilisation. On notera sur le dessin de la figure 14 le profil tourmenté de cette membrane, ses nombreuses expansions et le considérable développement de l’extrémité en massue ; la surface est ainsi notablement augmentée, et la sensibilité du système accrue. Cette sensibilité est telle que le corpuscule de Pacini peut répondre à un enfoncement de la peau de l’ordre de 0,5 à 1 μm.
Pour la genèse du potentiel d’action, on se reportera à l’article sensation, en notant qu’il a été établi avec certitude, dans le cas du corpuscule de Pacini, que le site générateur était constitué par le premier nœud de Ranvier (intracapsulaire).
Le codage dans le corpuscule de Pacini et le dôme de Iggo
On admet que le processus que nous venons de décrire s’applique, dans son principe, au fonctionnement des autres mécanorécepteurs cutanés. Mais, en ce qui concerne le codage, chaque dispositif possède ses caractéristiques propres, dont il tire sa spécificité. Nous le montrerons en comparant la possibilité de codage du corpuscule de Pacini à celle du dôme de Iggo, représenté sur la figure 6 et dont on voit la réponse à la figure 5.