



 Éd. 1971-1976
Éd. 1971-1976population
Ensemble des individus appartenant à la même espèce et occupant un espace déterminé à un moment donné.
La population animale
Les divers endroits favorables à son installation étant le plus souvent séparés entre eux par des discontinuités, une espèce est formée par un ensemble de populations isolées entre elles. Les Carpes d’un étang, les Escargots d’une haie, les Merles d’un parc en milieu urbain constituent des exemples de populations. Les divers individus d’une population étant capables de se reproduire entre eux, il existe un fond génétique commun ou pool de gènes répartis entre tous ses membres et dont la distribution est constamment modifiée au hasard des croisements. L’hétérogénéité génétique est la règle dans les populations naturelles. Sous l’action du milieu, des génotypes peuvent se trouver sélectionnés, ce qui conduit à la formation de races locales, surtout dans le cas de populations fermées n’entretenant pas d’échanges avec l’extérieur.
L’étude des populations, de leurs dimensions, de leurs structures, de leurs caractères généraux et de leur évolution constitue la démographie*, ou dynamique des populations. Les spécialistes de la démographie humaine ont joué un rôle pionnier, car ils ont disposé très tôt des données statistiques que les zoologistes ont encore bien du mal à obtenir. Ils ont mis au point les méthodes dont l’emploi a permis de faire progresser l’étude des populations animales.
Les caractéristiques des populations
La densité
La densité, c’est-à-dire le nombre d’individus par unité de surface ou de volume, est souvent difficile à évaluer. Des méthodes d’échantillonnage doivent être mises au point dans chaque cas (M. Lamotte et F. Bourlière, 1969, 1972). La difficulté principale dans l’estimation de la densité tient à la répartition non uniforme des animaux ; parfois la distribution se fait au hasard ; le plus souvent, elle est de type « contagieux » ou « par groupes ».
Le taux intrinsèque d’accroissement naturel
Le taux intrinsèque d’accroissement naturel r, par individu et dans des conditions de milieu données, est défini dans le cas d’une population à croissance exponentielle par la relation 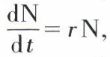 dans laquelle N est l’effectif et t le temps. On en déduit : Nt = N0ert, Nt étant l’effectif au temps t et N0 l’effectif au temps zéro. Le taux instantané d’accroissement naturel a été calculé chez l’Homme, puis chez certains Insectes et Rongeurs. Sa détermination nécessite de laborieux calculs. Elle permet de prévoir la vitesse de croissance des populations, au moins lorsque les conditions sont les plus favorables, et de comparer entre elles diverses espèces. La connaissance de r intervient dans plusieurs modèles mathématiques.
dans laquelle N est l’effectif et t le temps. On en déduit : Nt = N0ert, Nt étant l’effectif au temps t et N0 l’effectif au temps zéro. Le taux instantané d’accroissement naturel a été calculé chez l’Homme, puis chez certains Insectes et Rongeurs. Sa détermination nécessite de laborieux calculs. Elle permet de prévoir la vitesse de croissance des populations, au moins lorsque les conditions sont les plus favorables, et de comparer entre elles diverses espèces. La connaissance de r intervient dans plusieurs modèles mathématiques.
Table de mortalité
C’est la représentation du devenir d’un ensemble d’individus nés durant une même période de temps. L’établissement d’une telle table nécessite la connaissance de l’âge à la mort et des diverses causes de mortalité. L’emploi de ces tables est utile par exemple dans les recherches sur les Insectes ravageurs des forêts.
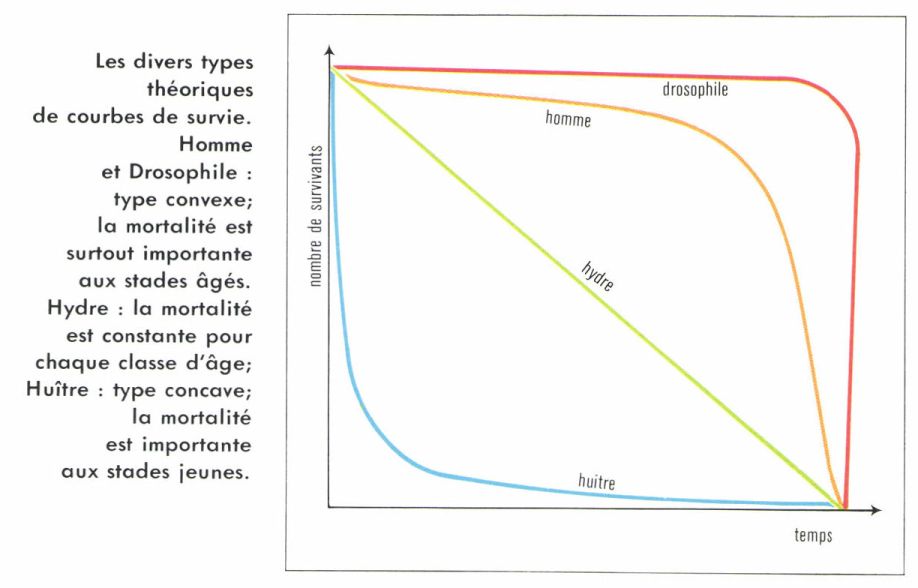
Courbe de survie
Cette courbe est tracée à partir de la table de survie, en plaçant le temps en abscisses et le nombre de survivants en ordonnées (échelle logarithmique). On ramène la population initiale à 1 000 individus afin d’avoir des courbes comparables.
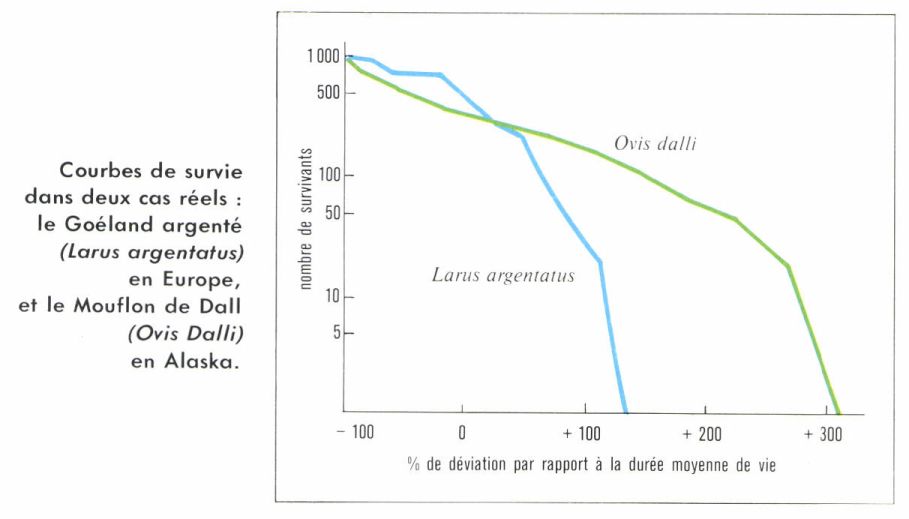
Pyramide des âges
Elle donne la structure d’une population par âge et par sexe.
La croissance des populations
Elle est due à deux phénomènes opposés : la natalité et la mortalité, auxquelles peuvent s’ajouter dans la nature l’émigration et l’immigration. La croissance exponentielle définie plus haut correspond au potentiel biotique de F. M. Chapman ; elle n’est évidemment pas possible indéfiniment dans un milieu limité. Vershulst, en 1845, a émis l’hypothèse de la croissance logistique des populations humaines ; cette loi a été appliquée aux populations animales disposant de ressources limitées, mais renouvelables, par R. Pearl (1925). On admet que la croissance des populations ralentit quand le nombre d’individus augmente, ce qui peut s’écrire en modifiant l’équation :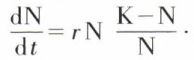
Le facteur correctif 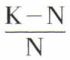 correspond à la résistance du milieu. On déduit de l’équation logistique
correspond à la résistance du milieu. On déduit de l’équation logistique 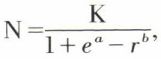 dans laquelle K est le nombre maximal d’individus (ou stock limite) pouvant vivre dans le milieu considéré ; K correspond à l’asymptote de la courbe. L’espace compris entre la courbe logistique et celle du potentiel biotique représente la résistance du milieu. Les valeurs a et b sont des constantes.
dans laquelle K est le nombre maximal d’individus (ou stock limite) pouvant vivre dans le milieu considéré ; K correspond à l’asymptote de la courbe. L’espace compris entre la courbe logistique et celle du potentiel biotique représente la résistance du milieu. Les valeurs a et b sont des constantes.
Plusieurs modèles expérimentaux ont montré la réalité de ce mode de croissance dans des cas simples tels que des élevages du Coléoptère Tribolium confusum ou de Protozoaires du genre Paramecium (expériences de Cause). Lorsque deux espèces qui ont la même niche écologique (c’est-à-dire les mêmes besoins) sont élevées ensemble, il y en a une qui est éliminée. C’est la loi de Gause, ou principe d’exclusion compétitive : deux espèces ayant la même niche écologique ne peuvent coexister. La mise en présence d’un prédateur et de sa proie aboutit à des variations cycliques d’abondance des deux espèces lorsque le milieu est assez complexe pour que des individus de l’espèce proie puissent échapper à leurs ennemis et se reproduire.
Les variations d’abondance des populations
Les fluctuations saisonnières sont fréquentes : la raréfaction des Insectes en hiver en est un bon exemple. Les fluctuations autour d’une valeur moyenne s’observent d’année en année chez beaucoup d’Oiseaux ; ces variations sont souvent en rapport avec des variations climatiques.
Les variations cycliques ayant une période de plusieurs années ont été analysées pour le Lièvre variable et le Lynx au Canada : les oscillations ont une période moyenne de 9,6 années, et le cycle d’abondance du Lièvre précède celui du Lynx de 1 à 2 ans. Les Lemmings ont des cycles de 4 ans ; la Tordeuse du Mélèze, en Engadine, un cycle de 10 ans.
