




physiologie (suite)
• Régulation de la circulation. Les quelques exemples exposés dans les paragraphes précédents montrent l’impérative nécessité d’une modulation des débits et pressions. Cette régulation s’effectue à deux niveaux : au niveau du cœur, par le ralentissement (nerf pneumogastrique ; fibres motrices) ou l’accélération (chaîne sympathique) du rythme cardiaque ; au niveau périphérique, par une vasoconstriction ou une vasodilatation principalement des artérioles, qui, en modifiant la résistance à l’écoulement (R), font varier le débit et la pression.
Physiologie respiratoire
La respiration* consiste en un échange d’O2 et de CO2 entre le milieu extérieur et l’organisme, le mécanisme de cet échange se résumant en termes de dissolution et de diffusion.
• Facteurs limitants. Le flux de diffusion d’O2 (I O2) d’un point A vers un point B dépend de plusieurs facteurs :
I O2 = α . P . S . ΔC ;
α = coefficient de solubilité de l’O2 dans l’eau (fonction de la température) ;
P = pression, qui dépend de la nature du gaz et du milieu organique, ainsi que de la longueur du trajet de diffusion (distance AB) ;
S = surface d’échange ;
ΔC = différence de concentration d’O2 entre A et B.
La teneur d’un liquide en gaz dissous s’exprime soit en termes de concentration (C), soit en termes de pression partielle (P) : C = α P ; α = coefficient de dissolution.
Les paramètres α, P et ΔC sont imposés par la nature de la source (20 p. 100 d’oxygène dans l’atmosphère, soit, au niveau de la mer, une pression partielle de 150 mm Hg), la nature et les besoins de l’organisme considéré et les propriétés physiques (dissolution) du gaz. Par contre, P et A peuvent être modifiés.
On conçoit aisément que, s’il existe une circulation qui draine l’O2 solubilisé au niveau de la surface d’échange, dont l’épaisseur peut être extrêmement fine (l’épaisseur de la paroi alvéolocapillaire est de l’ordre du micron), P devienne pratiquement négligeable. Mais un tel système fait surgir un nouvel élément : la nécessité d’un débit circulatoire suffisamment intense pour que l’apport d’oxygène, au niveau des tissus, corresponde à leurs besoins. La présence d’un transporteur (hémoglobine) capable de fixer des quantités relativement importantes d’hémoglobine permet de réduire de manière appréciable le débit circulatoire.
La surface d’échange est augmentée par la création d’organes spécialisés (poumon). Chez l’Homme, cette surface atteint 70 m2. L’existence d’une telle surface d’échange impose une nouvelle contrainte. Ses dimensions, sa fragilité, la nécessité qu’elle soit en contact avec une atmosphère saturée de vapeur d’eau (existence d’un film liquidien permettant la dissolution de l’O2) impliquent qu’elle soit pratiquement enclose ; d’où l’obligation d’une ventilation permettant le renouvellement de l’air pulmonaire (fig. 3).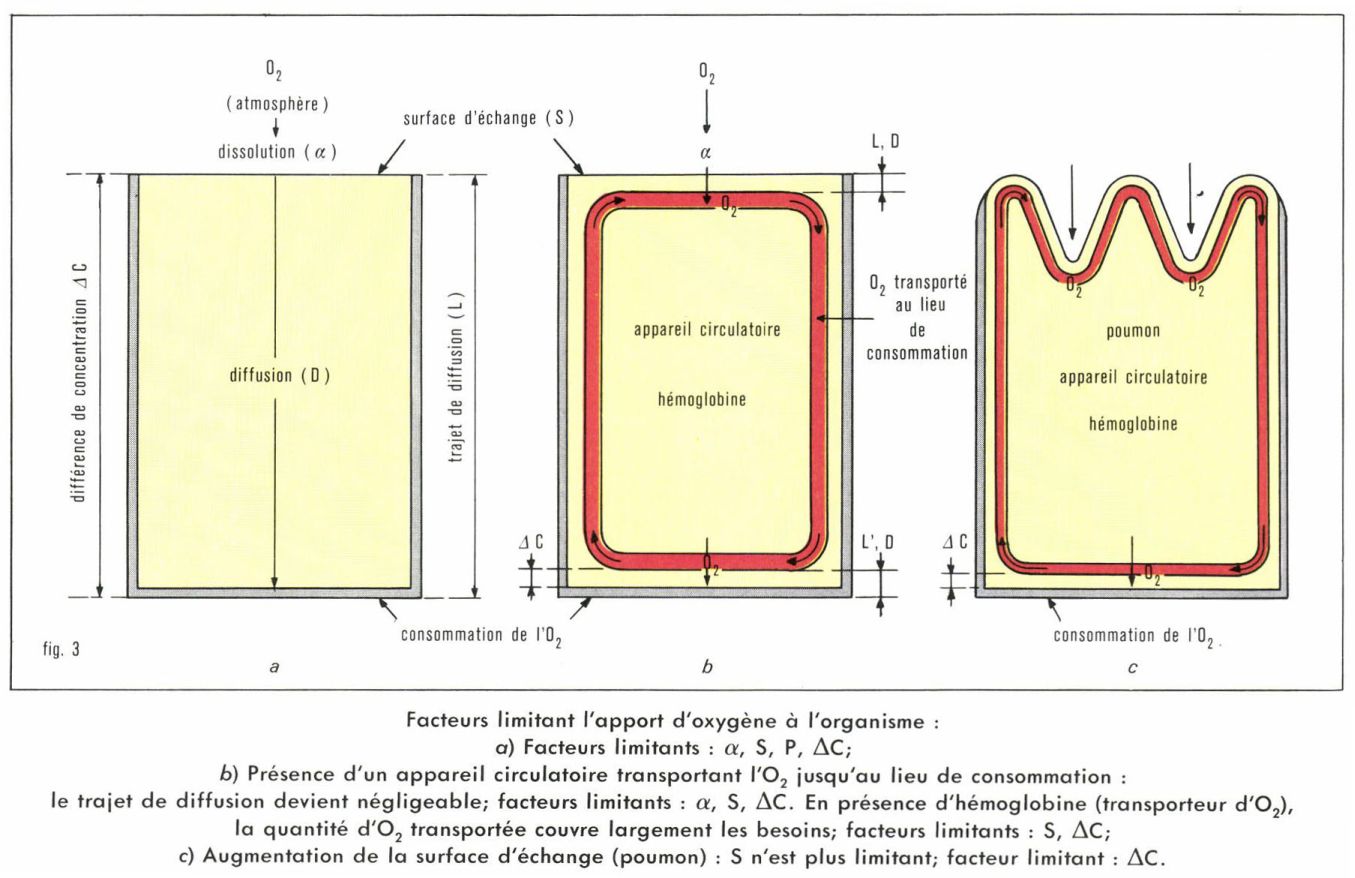
Les problèmes soulevés par la respiration sont donc : le mécanisme de la ventilation et sa régulation, les échanges alvéolocapillaires, le transport des gaz dans le sang, les échanges entre les capillaires et les cellules.
• Ventilation. Les poumons* sont enclos dans la cage thoracique et communiquent avec l’extérieur par l’intermédiaire de la trachée artère. Les poumons étant solidarisés (plèvres) à la cage thoracique, une augmentation de volume de cette dernière par élévation des côtes (muscles thoraciques) et abaissement (contraction) du diaphragme entraîne un appel d’air (inspiration). Ce type de ventilation est dit « à pression négative », par opposition à la ventilation « à pression positive », où l’air est introduit dans la bouche et propulsé dans les poumons (Amphibiens, Reptiles). Une fraction seulement (env. 1/3) de l’air inspiré atteint les alvéoles et participe aux échanges.
Les alvéoles peuvent être assimilés à une interface eau-air, ce qui implique l’existence d’une tension superficielle qui tend à faire refermer l’alvéole sur lui-même ; cette tension doit être équilibrée par la pression de l’air alvéolaire (cf. la loi de Laplace). S’il n’existait dans le film liquidien recouvrant la paroi alvéolaire des substances tensioactives (surfactants), un tel système serait instable. En effet, le volume des alvéoles étant variable, leur tension est donc variable ; par contre, la pression de l’air alvéolaire est la même pour tous, puisque les alvéoles communiquent entre eux. Dans ces conditions, les alvéoles à forte tension devraient se vider dans les alvéoles à faible tension ! Les surfactants ont la propriété d’abaisser la tension superficielle ; comme ils sont insolubles dans l’eau, leur concentration (par unité de surface) est d’autant plus grande que l’alvéole est plus petit, ce qui permet la stabilité du système alvéolaire. L’absence éventuelle de surfactants chez le nouveau-né gêne, sinon empêche, l’instauration de la ventilation (syndrome de détresse respiratoire).
• Échanges au niveau des tissus. La pression partielle de l’oxygène dans le sang artériel (Pα O2) est équilibrée avec celle de l’air alvéolaire : 100 mm Hg. La pression de l’oxygène dans les tissus est difficile à mesurer ; on peut l’estimer voisine de 40 mm Hg (Pτ O2). Pour qu’il y ait diffusion d’O2 du sang artériel vers les tissus, il est indispensable que Pα O2 soit supérieur à Pτ O2. Dans la mesure où la quantité de O2 disponible n’est pas un facteur limitant, compte tenu de la présence de l’hémoglobine, la Pα O2 peut avoir, sans inconvénients, des valeurs nettement inférieures à 100 mm Hg ; mais dès l’instant où Pα O2 = Pτ O2, il n’y a plus de diffusion possible. C’est à une altitude d’environ 6 000 m que la Pα O2 atteint la valeur limite de 40 mm Hg.
• Régulation de la ventilation. La théorie générale admise actuellement est celle des 3 facteurs : O2, CO2 et H+. Il faut une diminution importante de la Pα O2 (< 60 mm Hg) pour qu’apparaisse un accroissement de la ventilation (hyperventilation). Par contre, la moindre variation de la Pα CO2 entraîne des modifications ventilatoires (augmentations = hyperventilation ; diminution = hypoventilation). Il en est de même en ce qui concerne les ions H+ ; l’intervention de ces derniers dans la ventilation s’explique par le fait que leur concentration dans le sang dépend de la concentration en CO2 selon la réaction :
CO2 + H2O ⇋ CO3 H– + H+.