




assimilation (suite)
Du carbone minéral au carbone organique
Une grande partie des cellules autotrophes tirent l’énergie nécessaire à la conversion du C minéral (CO2) en C organique de l’énergie des radiations lumineuses émises par le soleil. L’assimilation est alors photosynthétique ou chlorophyllienne (v. photosynthèse). Certaines cellules autotrophes tirent leur énergie de réactions chimiques d’oxydation portant sur des composés minéraux. L’assimilation est alors chimiosynthétique.
Les premiers produits de l’assimilation du C sont des composés glucidiques simples (oses). En dérivent ultérieurement les glucides complexes, les acides gras et les lipides, les acides aminés et les protéines. Dans ce dernier cas s’y ajoute donc l’assimilation du N.
L’élaboration du C organique par chimiosynthèse est le privilège de certaines Bactéries, dont les plus remarquables sont les Bactéries de la nitrification : Nitrosomonas et Nitrosobacter. Les réactions d’oxydation génératrices d’énergie portant successivement sur NH4 (nitrosation) et NO2H (nitratation), leur importance découle en fait de leur participation au cycle d’assimilation du N. Le bilan en C organique des Bactéries chimiosynthétiques est négligeable devant le bilan des photosynthétiques.

Du carbone organique au carbone organique
Alors que, par sa forme gazeuse diffusible, le CO2 de l’atmosphère est à la disposition aisée des autotrophes immobiles que sont les végétaux pluricellulaires, il n’en est pas de même pour le C organique, qui est essentiellement sous forme solide fixée. Aussi les animaux, mobiles et actifs, sont-ils les plus aptes à utiliser en hétérotrophes le C organique préexistant.
Les cellules hétérotrophes doivent dégrader, à des stades divers, les molécules organiques qu’elles absorbent, puis réassembler de façon spécifique ces produits simplifiés afin de construire leurs propres molécules organiques. Les molécules simples résultant de la dégradation des divers composés carbonés sont au carrefour des réutilisations cellulaires pour les synthèses aussi bien des protéines que des glucides et des lipides. (V. métabolisme.)
De l’azote minéral à l’azote organique
Ce sont les formes non oxygénées : N2 moléculaire de l’atmosphère et NH3 des sels ammoniacaux, qui sont directement converties, par des systèmes distincts, en N organique. Les formes oxydées sont également utilisées, mais indirectement après réduction jusqu’au stade NH3. Les micro-organismes et les cellules végétales sont seules capables d’assimiler le N minéral. Hétérotrophes pour le C, les animaux le sont aussi pour le N.
De l’azote atmosphérique à l’azote organique
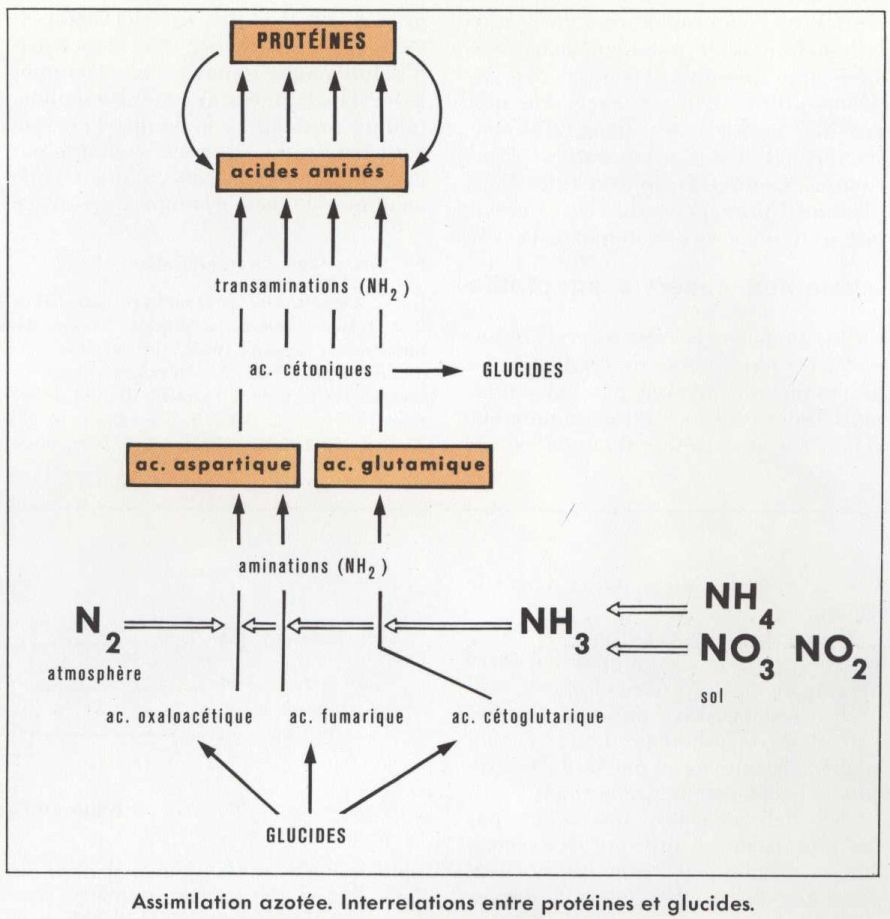
Malgré son abondance et sa disponibilité, l’assimilation du N2 de l’atmosphère est le privilège de quelques espèces seulement de Bactéries. Les unes sont libres, comme Azotobacter aérobie et Clostridium anaérobie. Les autres sont fixes et symbiotiques, comme Pavetta des feuilles de certaines plantes indiennes et surtout comme Rhizobium (ou Bacillus radicicola) des racines des Légumineuses. Signalons, outre les Bactéries, quelques Champignons inférieurs, libres ou symbiotiques. L’énergie nécessaire aux synthèses provient de l’oxydation des glucides, qui ont ainsi un rôle double : énergétique et chimique en tant que source du squelette carboné, accepteur du groupe aminé NH2. Les protéines résultent de l’association des acides aminés selon des agencements spécifiques.
De l’azote ammoniacal à l’azote organique
Les unicellulaires et les cellules végétales, dans leur ensemble, sont capables de transformer NH3 minéral en NH2 organique. Le groupe aminé est fixé sur des accepteurs carbonés spécifiques : d’une part, un acide insaturé, l’acide fumarique ; d’autre part, deux acides cétoniques, l’acide oxaloacétique et l’acide cétoglutarique. On obtient les deux premiers acides aminés, l’acide aspartique et l’acide glutamique, d’où dérivent les autres par transfert du groupe aminé, ou transamination.
D’un point de vue quantitatif, on peut estimer à près de 2 milliards de tonnes le N minéral transformé en N organique par an.
De l’azote organique à l’azote organique
Étant donné que les protéines d’une cellule ou d’un organisme donné sont spécifiques et différentes des protéines d’un autre organisme, le N organique subit des remaniements assurant cette spécificité. C’est au sein de chaque cellule et selon ses besoins structuraux que se déroulent les conversions éventuelles d’un acide aminé en un autre par transamination et l’assemblage des divers acides aminés en molécules de protéines. (V. métabolisme.)
Ces réactions, ainsi que l’importance des constituants glucidiques et lipidiques dont l’oxydation est à l’origine de l’énergie nécessaire aux synthèses, illustrent les interrelations multiples entre assimilation azotée et assimilation carbonée.
Déterminisme de la spécificité des synthèses
Par le jeu incessant de l’élaboration et de la destruction des composés organiques, ces composés subissent un renouvellement constant, auquel n’échappent pas d’ailleurs les composés minéraux. Dans la biosphère, les éléments C, H, O et N sont en état de flux permanent, plus ou moins rapide, au sein des divers composés cellulaires.
On peut se demander quel est le déterminisme orientant spécifiquement les synthèses de matière vivante. Étant donné que les acides nucléiques (ADN) sont le support potentiel de tous les caractères héréditaires, ils sont les facteurs premiers de cette spécificité. Si la séquence ADN → ARN → protéines met bien en évidence la spécificité des protéines, elle n’explique pas à première vue celle des glucides et des lipides. Toutes les synthèses biologiques en jeu dans le phénomène de l’assimilation étant dirigées par les enzymes, catalyseurs spécifiques, et les enzymes étant de nature protéique, les diverses spécificités d’orientation relèvent cependant toutes de la spécificité des protéines codées par les acides nucléiques.

M. P.