nerveux (système) (suite)
L’axone et ses gaines
L’axone est la partie allongée du neurone, qui assure la conduction des influx nerveux. Il est constitué d’une membrane cytoplasmique, ou axolemme, et de cytoplasme, ou axoplasme, renfermant quelques mitochondries et surtout des neurofilaments nombreux. Ces derniers ne jouent pas de rôle conducteur vis-à-vis de l’influx nerveux, comme on le croyait autrefois, mais constituent un squelette élémentaire. L’axone est très rarement nu ; il est entouré de cellules névrogliques (cellules de Schwann dans le système nerveux périphérique, oligodendrocytes dans le système nerveux central) ; on appelle mésaxone l’affrontement membranaire des deux bords de la cellule névrotique qui a entouré l’axone. On parle de fibre amyélinique quand le mésaxone est court et de fibre myélinisée quand le mésaxone s’enroule en spirale autour de l’axone.
Dans le cas des fibres amyéliniques, on trouve en général plusieurs axones qu’enveloppe et isole les uns des autres une cellule de Schwann. Les cellules de Schwann se succèdent en chapelets le long de ces fibres. On trouve en moyenne de 4 à 12 axones ainsi enveloppés, mais ce nombre va jusqu’à la centaine dans le nerf olfactif des Vertébrés. Il existe également des fibres amyéliniques enveloppées individuellement par un chapelet de cellules de Schwann ; ces fibres annoncent les fibres myélinisées, très abondantes dans la substance blanche, à laquelle elles donnent sa couleur. Dans une fibre myélinisée, chaque cellule de Schwann vient entourer un seul axone, puis le mésaxone ainsi formé s’enroule en formant des spirales externe et interne. Les membranes s’accolent, et leur ensemble constitue une masse très réfringente. Au niveau où deux cellules de Schwann sont accolées, la gaine de myéline s’interrompt, et l’axolemme est au contact du milieu extérieur : c’est l’étranglement de Ranvier. Les fibres amyéliniques sont en général fines (diamètre égal ou inférieur à 1 μ, et les fibres myélinisées plus épaisses.
La synapse
On appelle synapse la zone d’accolement d’un bouton terminal avec la région dendritique d’un autre neurone ou avec la membrane d’un effecteur. Dans le cas des fibres musculaires, le contact synaptique prend le nom de plaque motrice. Dans le cas des synapses interneuroniques, la membrane postsynaptique peut être différenciée morphologiquement (on l’appelle alors épine dendritique, bien que sa forme soit le plus souvent arrondie). Les deux membranes pré- et postsynaptique, appartenant à deux neurones distincts, sont séparées par un espace synaptique d’environ 0,02 μ d’épaisseur, que seul le microscope électronique met en évidence. Le bouton terminal est un petit renflement qui prolonge l’axoplasme et renferme les mêmes organites cellulaires, et en particulier un grand nombre de mitochondries. On y trouve également des vésicules, dites synaptiques, de 0,02 à 0,05 μ de diamètre.
La physiologie du neurone
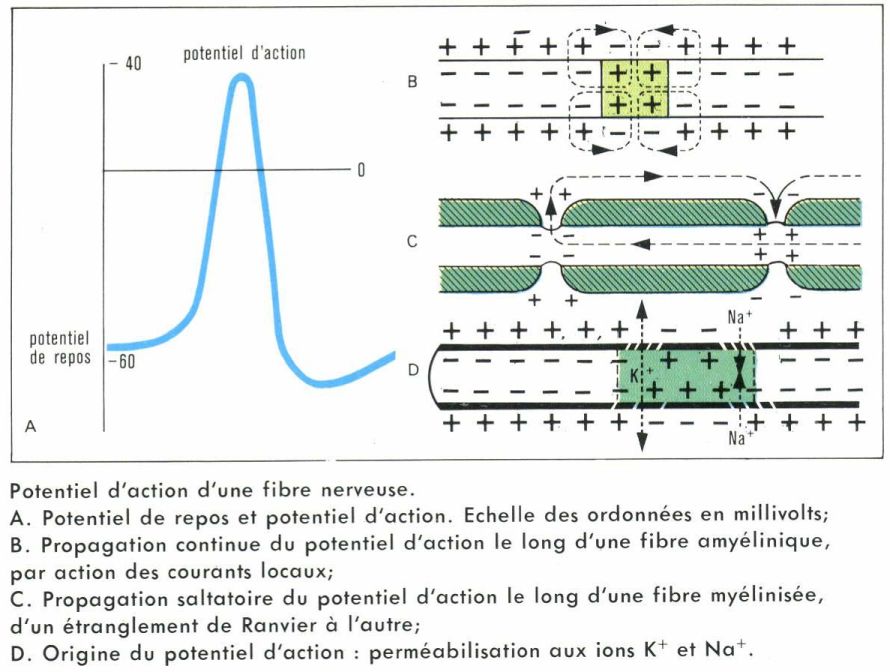
Le potentiel d’action
La membrane cytoplasmique du neurone est polarisée électriquement ; si l’on introduit une micro-électrode à l’intérieur de l’axoplasme, l’autre étant dans le milieu extracellulaire, on constate que l’intérieur de la membrane est négatif par rapport à l’extérieur. Cette différence de potentiel, dite potentiel de repos, est de l’ordre de 70 mV. Toute stimulation efficace portée sur un récepteur sensoriel ou sur un neurone modifie localement ce potentiel de repos ; il se produit une dépolarisation dont l’amplitude dépend de l’intensité de la stimulation qui a été portée. Pour une intensité de stimulation supérieure à un certain seuil, on observe la naissance d’un potentiel d’action qui a comme particularités :
— d’avoir une amplitude indépendante de l’intensité de l’excitation, pourvu que cette dernière soit supraliminaire ;
— d’être transmissible le long de l’axone sans perdre de son amplitude.
On explique la genèse de ce potentiel d’action par le fait que la membrane du neurone, à l’endroit de l’excitation, est brusquement rendue perméable aux ions Na+ et K+. Ce potentiel d’action dure en moyenne un millième de seconde. Il est suivi d’une période, également très courte, au cours de laquelle la membrane est inexcitable ; c’est la période réfractaire absolue.
Le potentiel d’action correspond à l’entrée brutale d’ions Na+ ainsi qu’à la sortie d’ions K+. La membrane retrouve son excitabilité en rétablissant, par un transport actif, coûteux en énergie, les différences de concentration de ces ions (ainsi que des ions Cl– et d’autres anions) de part et d’autre d’elle.
La conduction du potentiel d’action
Dans la fibre myélinisée, les seuls endroits où les flux ioniques sont possibles sont les étranglements de Ranvier. Le potentiel d’action qui existe au niveau d’un de ces étranglements provoque dans l’étranglement voisin, par transfert de charges, une dépolarisation supraliminaire qui donne naissance à son tour à un nouveau potentiel d’action ; on dit que la conduction est saltatoire — car la myéline joue le rôle d’isolant électrique —, et le potentiel d’action se régénère au niveau de chaque étranglement de Ranvier, ce qui explique qu’il soit conduit sans décrément (sans perdre de son amplitude). Le phénomène est le même dans la fibre amyélinique : les courants locaux provoqués par le potentiel d’action en un point dépolarisent la membrane proche avec une intensité supraliminaire.
La vitesse de conduction est directement proportionnelle au diamètre de la fibre myélinisée ; elle est proportionnelle à la racine carrée de ce diamètre pour les fibres amyéliniques. Elle varie de quelques millimètres à une centaine de mètres par seconde. Elle est évidemment fonction de la vitesse des courants locaux : il suffit de plonger une fibre nerveuse dans un milieu moins conducteur pour observer un ralentissement de cette vitesse.
La conduction du potentiel d’action le long de l’axone, à partir d’un point de stimulation, a lieu indifféremment dans les deux sens, mais un seul sens est fonctionnellement efficace, celui qui va des régions dendritiques vers les arborisations terminales, où le potentiel d’action peut être transmis, à travers la synapse, à un autre neurone ou à un effecteur.