




motricité (suite)
• Organisation fonctionnelle du système thalamo-strié
Le thalamus distribue les afférences. Le striatum les intègre. Le pallidum a un rôle d’effecteur. On décrit ainsi trois circuits fonctionnels.
Le circuit pallido-thalamo-striatal. Les voies effectrices du pallidum, qui constituent l’anse lenticulaire, sont en connexion avec les noyaux ventraux latéraux et le noyau ventral antérieur du thalamus. Ces structures sont elles-mêmes en relation avec le striatum. Ce circuit entre en relation avec les aires corticales motrices et contrôle ainsi les influx efférents moteurs qui partent du cortex.
Le circuit pallido-sous-thalamique. Les fibres de l’anse lenticulaire entrent en relation avec les noyaux hypothalamiques, sous-thalamiques et la réticulée mésencéphalique, et exercent ainsi une influence sur les motoneurones gamma statiques (v. moelle épinière).
Le circuit nigro-striatal. Le locus niger (autre noyau gris central du cerveau) et le striatum sont étroitement unis par une boucle neuronale fermée à fonctionnement dopaminergique (dont le médiateur chimique est la dopamine [v. schéma]).
La stimulation du locus niger déclenche une libération de dopamine au niveau du striatum homolatéral ; une lésion du locus niger est suivie d’une chute du taux de dopamine au niveau du striatum.
Dans la maladie de Parkinson, on observe de façon presque constante une altération des neurones du locus niger ; or, la dopamine est présente normalement à un taux élevé au niveau du locus niger ; chez les parkinsoniens et chez les sujets traités pendant longtemps par la réserpine et les phénothiazines, elle est à un taux très bas.
On peut donc admettre que le syndrome parkinsonien (tremblement, hypertonie, akinésie) résulte d’un mauvais fonctionnement de la boucle nigro-striée et peut donc, dans certains cas, être corrigée par l’administration de dopamine.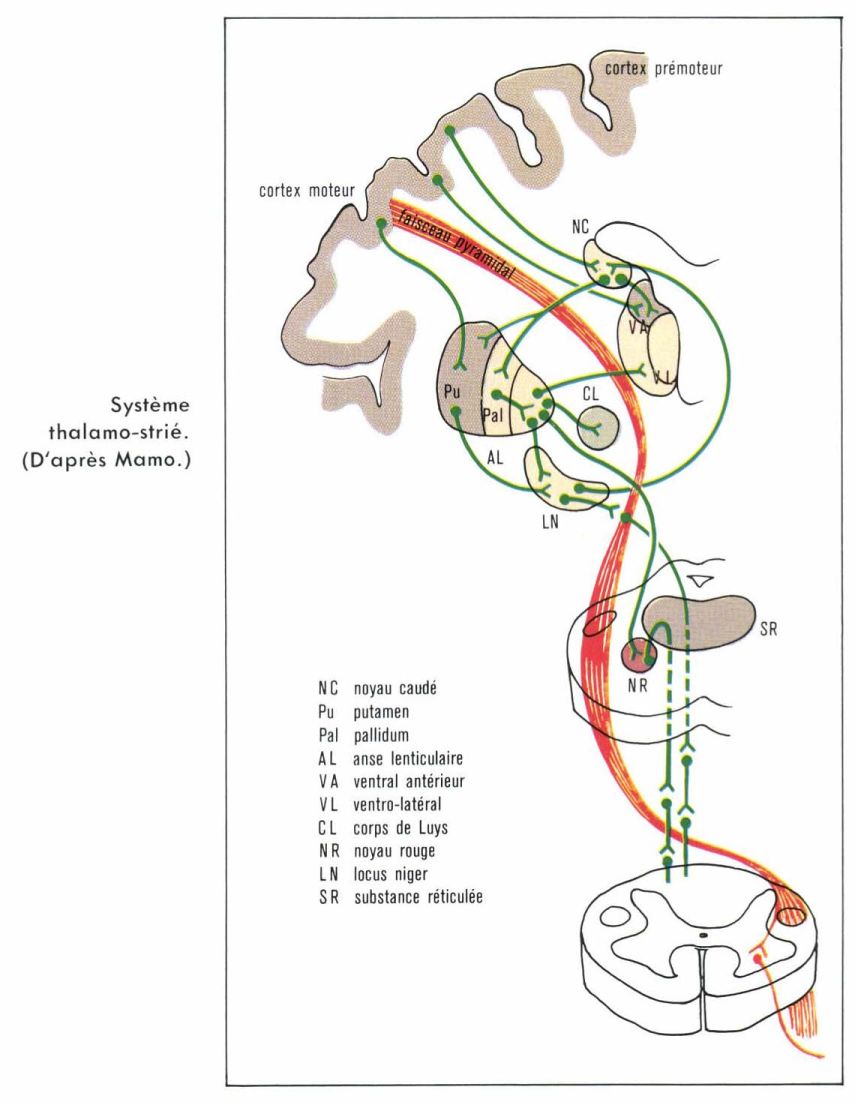
• Rôle des noyaux gris centraux
Les noyaux gris centraux exercent une double action sur le contrôle du mouvement.
Ils achèvent l’acquisition de la station debout en contrôlant les informations venues du tronc cérébral (afférences vestibulaires et visuelles).
Ils modulent les activités toniques fondamentales.
Les mécanismes de redressement. Ils dépendent du tronc cérébral, mais l’achèvement de la station debout nécessite l’intégration des afférences vestibulaires et visuelles au niveau thalamo-pallidal.
Lors d’une lésion sous-thalamique ou bilatérale du pallidum, on observe une dystonie en flexion due à la libération des dispositifs réflexes du tronc cérébral.
L’adaptation posturale. Le circuit thalamo-strié est responsable du maintien de la station debout. Lors de la marche, par exemple, le déplacement de l’axe du corps par rapport à la verticale demande une adaptation en série du tonus musculaire pour maintenir l’équilibre. C’est grâce à l’intégration des afférences proprioceptives vestibulaires, cutanées, visuelles par le centre médian du thalamus que ce phénomène peut se réaliser. Le pallidum en est l’effecteur.
L’adaptation des mouvements segmentaires au monde extérieur dépend aussi des noyaux gris centraux.
Le Chat sans cortex se lèche, se nourrit spontanément, a un comportement sexuel. Chez les Primates, l’évaluation de cette participation est difficile.
La stimulation du cortex extra-pyramidal déclenche des effets moteurs controlatéraux de caractère global et grossier. L’exérèse de ces régions produit une réduction de l’activité spontanée ou l’exagération des comportements moteurs fondamentaux.
La préhension pathologique, sous la dépendance du lobe pariétal, est libérée par des lésions du lobe frontal. La réaction d’évitement, sous la dépendance du lobe frontal, est libérée par des destructions du lobe temporal. En résumé, les noyaux gris centraux et le cortex extra-pyramidal sont responsables des adaptations spatiales fines de la réponse motrice en fonction de la stimulation.
Il semble exister au niveau des noyaux gris centraux une certaine spécialisation.
1. Le noyau caudé joue un rôle important dans la réalisation des déséquilibres contrôlés nécessaires à l’exécution de la marche.
2. Le putamen intervient sur la mobilité d’attitude.
3. L’action inhibitrice du locus niger est indispensable pour établir l’équilibre entre les afférences toniques et phasiques au sein du striatum.
4. Le corps de Luys est indispensable pour limiter certaines réactions phasiques du pallidum aux afférences vestibulaires.
Les structures corticales (cortex cérébral)
Le cortex cérébral exerce son contrôle sur tous les étages du névraxe en empruntant des voies nombreuses et variées issues de zones bien définies. La plus importante et la mieux connue est la voie pyramidale. Par des expériences de stimulation électrique et d’ablation, on a pu identifier des zones corticales dont l’excitation provoque des mouvements systématisés.
• L’aire motrice rolandique
Elle occupe chez le Singe et l’Homme la circonvolution frontale ascendante située en avant de la scissure de Rolando. La stimulation électrique de cette région a permis de préciser l’existence d’une représentation régionale des diverses parties controlatérales du corps, selon la disposition d’un hémi-corps renversé. L’importance en surface corticale donnée aux pattes antérieures et postérieures chez le Singe, la large représentation de la main et du pouce chez l’Homme comparativement au tronc soulignent le rôle de cette région du cortex dans la finesse du contrôle moteur de ces segments.
Schématiquement, lorsqu’on fait une coupe du cortex, on voit que l’agencement des corps cellulaires est tel que l’on peut distinguer six couches (couche moléculaire, couche granulaire externe, couche pyramidale externe, couche granulaire interne, couche pyramidale interne, couche fusiforme), en allant de la superficie à la profondeur.
Cela permet de diviser l’écorce en deux grands territoires : un allocortex, de structure primitive, où seules deux couches sont individualisées, et un isocortex, où existent nettement les six couches.
Lorsque les couches granulaires prédominent, le cortex est spécialisé dans la réception sensitive ; lorsque les couches pyramidales l’emportent, il est dit « moteur » ; c’est le cas du cortex rolandique.