




métamorphoses (suite)
Hétérométaboles
Ils sont caractérisés par des métamorphoses progressives : la larve diffère peu de l’adulte, et les mues qu’elle subit la rapprochent peu à peu de la forme imaginale qu’elle atteint directement, sans stade nymphal immobile ; les fourreaux alaires (ptérothèques) apparaissent sur le thorax au cours des derniers stades larvaires, mais les ailes ne se déploient et ne deviennent fonctionnelles qu’à la mue imaginale, comme d’ailleurs l’appareil génital.
Les Dictyoptères (Blattes, Mantes), les Isoptères (Termites), les Chéleutoptères (Phasmes), les Orthoptères (Criquets), les Dermaptères (Perce-Oreilles), les Psocoptères (Psoques), les Mallophages et les Anoploures (Poux), les Hétéroptères (Punaises) et les Homoptères (Cigales) montrent un tel développement ; on les qualifie souvent de « paurométaboles » pour les distinguer de deux ordres, les Odonates (Libellules) et les Plécoptères (Perles), dont les larves mènent une vie aquatique et ont des organes particuliers qui disparaissent à la dernière mue (« hémimétaboles »).
Chez les Thysanoptères, les deux derniers stades larvaires, appelés pronymphe et nymphe, se déroulent dans des conditions qui rappellent les vraies nymphes des holométaboles : l’animal ne se nourrit pas et reste inactif, les pièces buccales et les antennes s’atrophient ainsi que certains muscles ; cependant, larves et adultes sont morphologiquement très proches. On qualifie ces Insectes de « remétaboles ».
Les Cochenilles femelles passent sans discontinuité de la larve à l’adulte ; mais le mâle a un développement plus complexe, comportant un stade de pupe avant la dernière mue. Les deux sexes d’une même espèce ont donc des métamorphoses très différentes ; c’est une raison pour ne pas faire du type de développement un caractère systématique sûr. Avec leurs larves aquatiques et leurs adultes aériens, les Ephémères se rapprochent des hémimétaboles ; mais la mue qui termine la vie larvaire donne un Insecte ailé (subimago) qui subit une nouvelle mue avant de libérer l’imago. L’existence de ce stade intermédiaire actif entre la larve et l’adulte est un cas unique chez les Insectes ; on le considère comme un caractère archaïque.
Amétaboles
Les Aptérygotes, les plus primitifs des Insectes, sont dépourvus d’ailes ; seules la taille plus élevée et la possibilité de reproduction permettent de distinguer l’adulte du jeune. Chez les Protoures, cependant, le nombre des segments abdominaux passe de 9 à l’éclosion à 12 chez l’adulte.
Déterminisme de la métamorphose
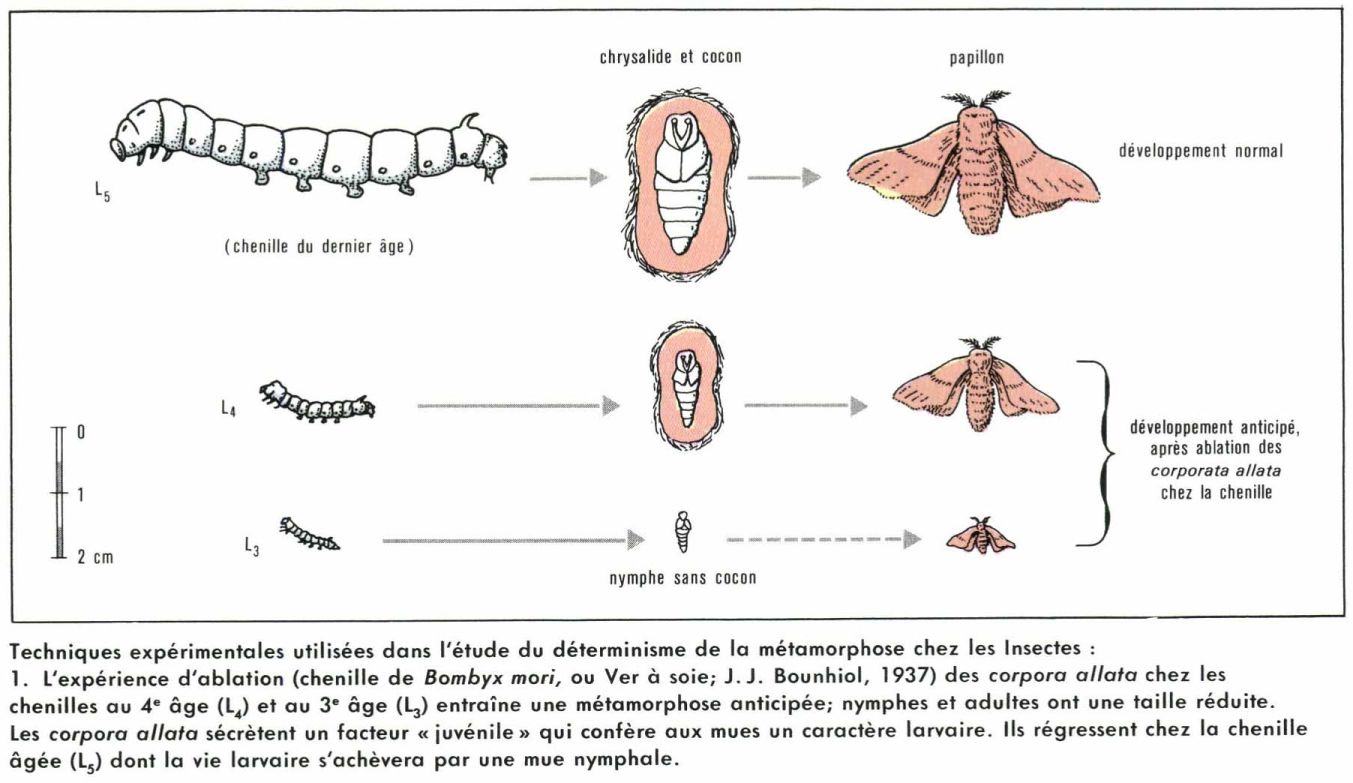
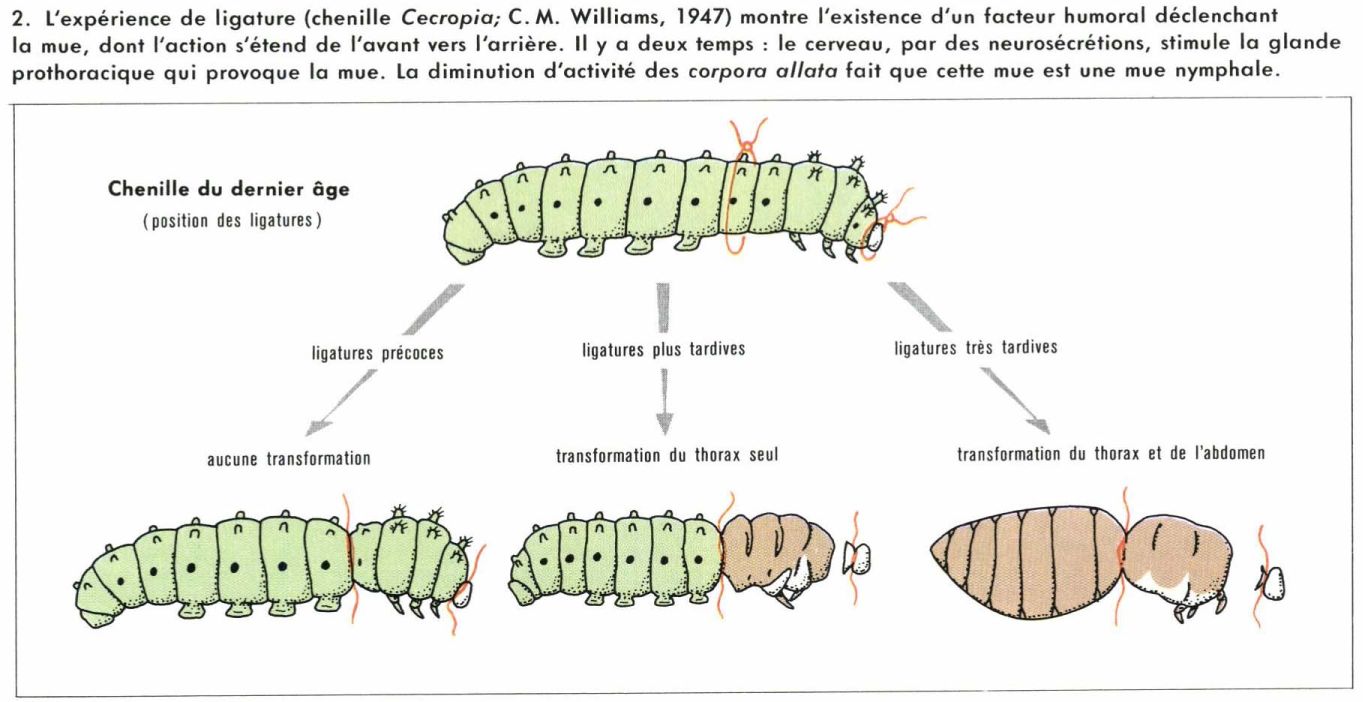
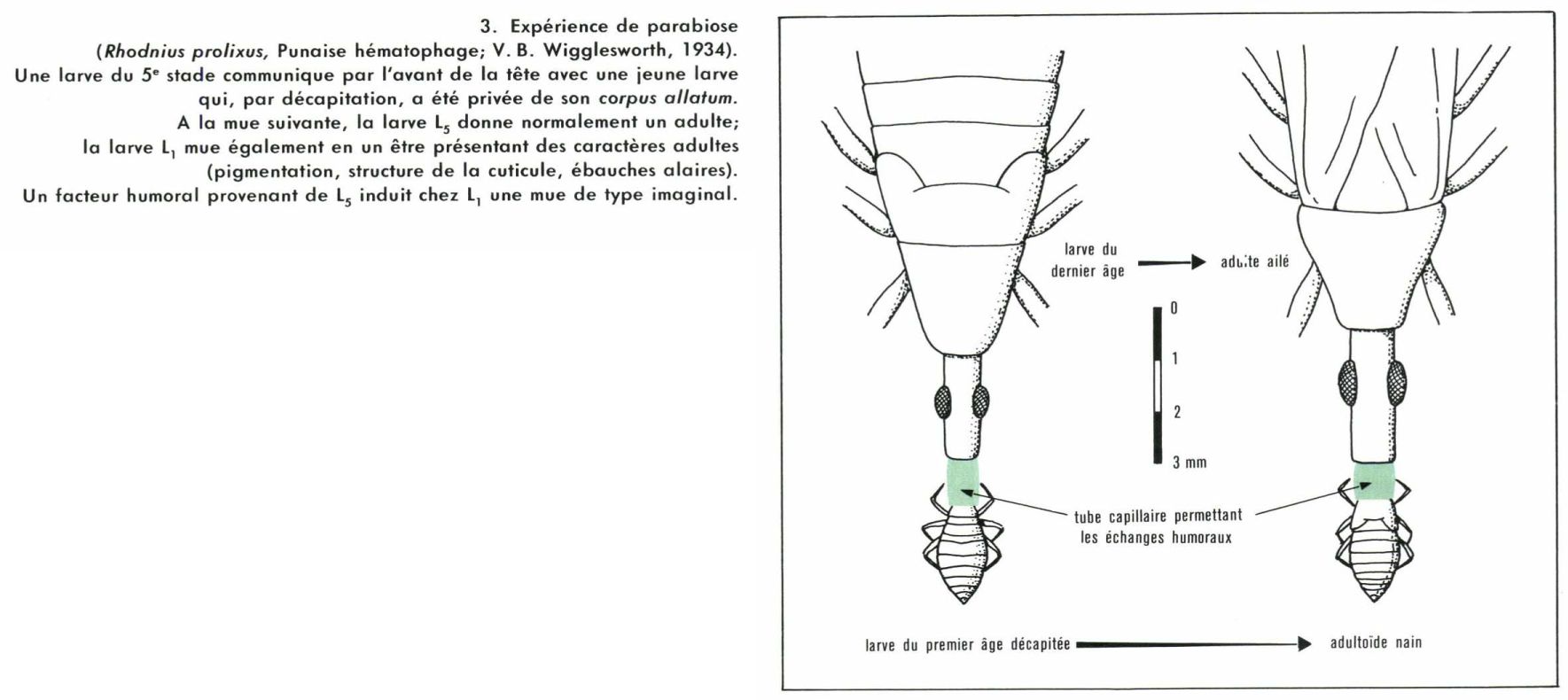
Mues et métamorphoses sont deux phénomènes indissociables dans le développement des Insectes. Ils sont sous la dépendance d’actions hormonales, qu’on peut ainsi résumer : chaque mue est contrôlée par une hormone appelée ecdysone ; si une autre hormone, dite « hormone juvénile », agit en même temps, la mue est du type larvaire ; si l’ecdysone agit seule, la mue est du type imaginal. Le déroulement normal de la métamorphose implique donc l’intervention simultanée des deux facteurs pendant la vie larvaire et la disparition plus ou moins rapide de l’hormone juvénile vers la fin. Bien entendu, le rythme et les lieux de sécrétion varient sensiblement d’un groupe à l’autre ; de nombreuses recherche expérimentales ont été entreprises pour préciser les modalités d’action de ces facteurs ; la petitesse des animaux exige une habileté exceptionnelle pour utiliser les techniques classiques de l’endocrinologie : destructions de glandes, implantations, ligatures, parabioses. Quelques résultats, retenus parmi les plus significatifs, serviront d’exemples.
Travaillant dès 1934 sur la Punaise hématophage Rhodnius prolixus, V. B. Wigglesworth a localisé l’origine de l’hormone juvénile dans les corpora allata situés en arrière du cerveau, près de l’œsophage ; ils s’arrêtent de fonctionner au cinquième stade, et la mue suivante fait apparaître l’imago. Quant à l’hormone de mue, elle provient de la glande prothoracique, dont l’activité est elle-même réglée par les cellules neurosécrétrices du cerveau.
En greffant des corpora allata provenant de jeunes Phasmes (Carausius morosus) à des larves âgées, Otto Pflugfelder provoque chez celles-ci des mues supplémentaires (jusqu’à cinq) et prolonge ainsi la métamorphose.
Les Lépidoptères ont été l’objet de multiples recherches montrant, chez ces holométaboles, l’intervention de mécanismes complexes ; des équilibres subtils, en évolution constante, règlent chaque étape du développement, même les plus stables en apparence, comme le stade imaginal. Les cellules neurosécrétrices de la portion médiane du cerveau antérieur (pars intercerebralis) libèrent un facteur qui, par l’intermédiaire probable des corpora cardiaca, agit sur les glandes prothoraciques ; minuscules organes ramifiés situés contre les trachées, celles-ci émettent l’hormone de mue (ecdysone) [travaux de S. Fukuda sur le ver à soie, en 1940] ; cette hormone a pu être isolée en 1953. Dès 1937, J. J. Bounhiol étudiait, sur le même ver à soie, le rôle des corpora allata et montrait que leur ablation entraînait une métamorphose anticipée ; l’hormone juvénile qu’ils produisent a pu être extraite de l’abdomen de Papillons Platysamia cecropia.
L’anneau de Weismann qui entoure l’aorte antérieure des larves de Diptères supérieurs réunit trois glandes séparées dans les autres groupes : corpora allata, corpora cardiaca, glande thoracique.
Le développement de nombreux Insectes peut être entrecoupé d’arrêts d’origine génétique ou déterminés par des facteurs externes, en particulier des variations de température (diapause), comme C. M. Williams l’a montré, dès 1947, sur un Papillon (Cecropia) à longue diapause nymphale ; cet état correspond à une déficience hormonale et peut être rompu par un traitement approprié.

Arachnides
Les Acariens montrent des métamorphoses compliquées, où se succèdent des stades différents par leur morphologie et leur biologie ; on compte six formes fondamentales (sept avec la prélarve incluse dans la membrane de l’œuf), mais parfois elles se télescopent et restent incluses les unes dans les autres. À l’éclosion, la larve n’a que six pattes ; elle subit une dédifférenciation complète avant de se transformer en protonymphe à quatre paires de pattes ; les stades suivants (deutonymphe, tritonymphe, imago) sont séparés par des mues où des phénomènes d’histolyse et d’histogenèse s’observent souvent.