




toucher ou tact (suite)
Ces descriptions résultaient d’études faites avec les moyens de la microscopie optique, et l’on ne disposait, au plan fonctionnel, que des données obtenues chez l’Homme par les méthodes expérimentales. Rapprocher les deux ordres de résultats était pour le moins aventureux ; on s’y risqua cependant et l’on peut, de nos jours encore, lire dans certains manuels que le corpuscule de Meissner est le récepteur du tact superficiel, celui de Pacini de la pression profonde, celui de Krause du froid et celui de Ruffini du chaud. Nous verrons qu’en ce qui concerne les trois derniers l’affirmation est radicalement erronée ; quant au corpuscule de Meissner, s’il est bien récepteur de pression, il faut ajouter que des régions cutanées peuvent être sensibles à la pression (légère ou profonde) bien que ne possédant aucun corpuscule. Les recherches contemporaines — bénéficiant, pour l’observation biologique, du microscope électronique et, pour l’étude fonctionnelle, des moyens de la micro-électrophysiologie — peuvent aborder ces problèmes avec plus de succès ; on ne saurait cependant prétendre que toutes les obscurités et tous les malentendus sont aujourd’hui dissipés.
La recherche des spécificités fonctionnelles
Si, chez un Rat, un Chat, un Singe, on introduit une micro-électrode dans un rameau sensitif cutané, on peut recueillir les réponses (potentiels d’action) d’une seule des fibres et déterminer, en exerçant des stimulations sur la peau, le mode de stimulation susceptible de déclencher l’activité de cette fibre. Une telle méthode permet de mettre en relation un certain type de fibre et une certaine modalité stimulatrice. Mais, pour que l’étude soit complète, il faut encore identifier la terminaison réceptrice de cette fibre. On y est parvenu dans certains cas, mais il reste encore à faire dans ce domaine. Nous ne rapporterons ici que les exemples les plus favorables, pour lesquels ne subsistent pas trop d’inconnues, et nous les classerons d’après la nature des stimuli efficaces en trois catégories : mécanorécepteurs, thermorécepteurs, nocicepteurs (c’est-à-dire récepteurs des stimulations douloureuses).
Mécanorécepteurs
• Récepteurs statodynamiques de pression. La figure 5 donne les réponses de deux types de ces récepteurs à un enfoncement de la peau de 0,5 mm (chez le Chat) maintenu pendant plusieurs dizaines de secondes. Pour le type I, la fibre, en l’absence de stimulation, a une activité quasi nulle. L’application de la stimulation provoque une décharge initiale importante dont la fréquence décroît rapidement, puis se maintient tant que la peau reste déformée. Pour le type II, en l’absence de stimulation, la fibre est déjà active et sa fréquence de repos (une vingtaine d’influx à la seconde) s’accroît transitoirement lorsqu’on déforme la peau, mais moins que dans le type I. On observe ensuite une décharge maintenue, et, lorsque cesse la stimulation, le retour à la fréquence de repos est précédé d’une diminution transitoire d’activité. Ces deux récepteurs se distinguent encore par le fait que le premier est plus sensible à la dynamique de la stimulation, c’est-à-dire à la vitesse avec laquelle on applique la déformation : si l’on déforme plus vite, la fréquence de la réponse est plus élevée ; le second est plus sensible au caractère statique, c’est-à-dire à l’amplitude de la déformation. La mesure des vitesses de conduction de l’influx sur les fibres a permis de préciser que, dans le cas du type I, il s’agit de fibres nerveuses dites « Aα » rapides, 55 à 75 m/s, ce qui correspond à un diamètre de 9 à 12,5 μm environ, et, dans le cas du type II, de fibres légèrement plus lentes (diamètre de 7,5 à 11 μm). Prélevant la portion de peau sur laquelle s’exerçait la stimulation, on a pu ensuite montrer que le récepteur de type I était constitué par un dôme de Iggo. Un tel dôme (fig. 6) accompagnait généralement un poil (fig. 7) et devait être situé en avant du poil chez le Chat et en arrière chez les autres Mammifères jusque-là étudiés (le caractère systématique de cette disposition peut être important pour la perception de la direction des déplacements à la surface de la peau). Situé dans la couche des cellules épidermiques basales, il est formé par une grappe de complexes de Merkel (disque + cellule), et le microscope électronique révèle, entre le disque nerveux et la cellule accompagnatrice, des images de type synaptoïde telles que l’on se demande si les cellules de Merkel ne joueraient pas, plutôt qu’un rôle passif de soutien, un rôle sensoriel actif.
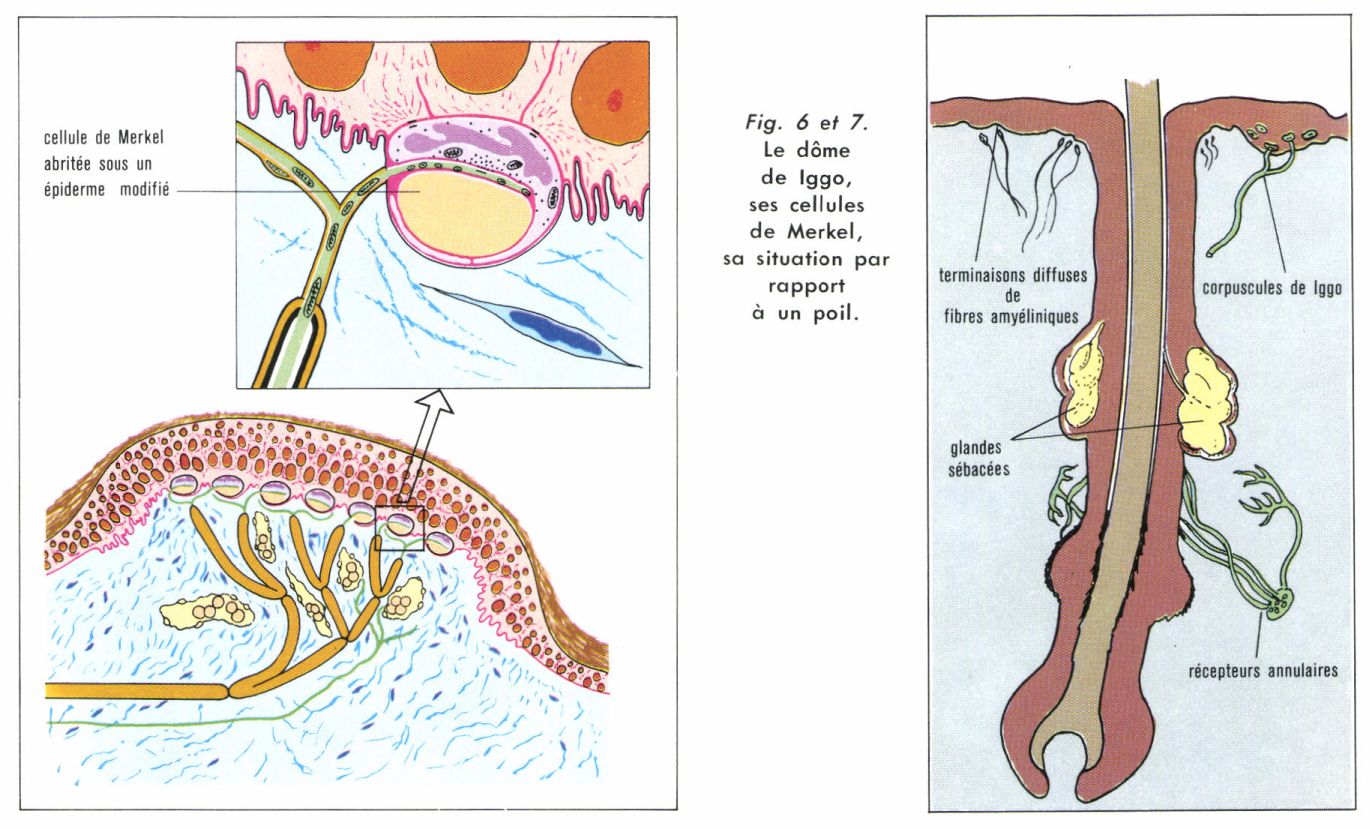
Quant aux récepteurs de type II, il semble qu’ils ne sont autres que des corpuscules de Ruffini (fig. 8). La présence de fibres conjonctiles à l’intérieur de ces corpuscules permettait de leur supposer un rôle mécano-récepteur qui se trouve ainsi confirmé. Dans les deux cas, les aires réceptrices sont très réduites et correspondent bien aux « points » des anciens auteurs.
• Récepteurs dynamiques (associés aux poils). On voit, figure 7, le schéma d’un cil de Macaque avec son innervation. Deux systèmes de terminaisons nerveuses lancéolées entourent la gaine au-dessous des glandes sébacées : des terminaisons orientées parallèlement au poil forment une sorte de palissade circulaire, et, en arrière de cette palissade, se trouvent des terminaisons orientées perpendiculairement à l’axe du poil. On peut présumer que la palissade réagit à de très faibles déplacements du poil, tandis que le second système n’intervient que lorsque les déplacements sont plus considérables. Ces terminaisons lancéolées sont celles de fibres rapides du type A, fibres qui répondent pendant que l’on incline le poil et avec une fréquence d’autant plus grande que le mouvement est plus rapide. Dès que le mouvement s’arrête, le poil restant incliné, l’activité cesse. Nous avons affaire là à un récepteur dynamique typique. On réussit, dans certains cas, à mettre en évidence une composante statique, mais il faut alors une amplitude de mouvement beaucoup plus grande. On pense que les terminaisons palissadiques sont responsables de la réponse dynamique, et les autres de la réponse statique, lorsqu’elle existe.
Dans le cas particulier des vibrisses, le poil, entouré d’un important sinus veineux, possède un équipement nerveux beaucoup plus riche : terminaisons lancéolées entourant le poil en anneau en plus des terminaisons précédentes : complexes de Merkel et terminaisons encapsulées entre le poil et le sinus.