




audition (suite)
Les stimulations sonores, captées par l’oreille, sont transmises par le tympan et les osselets à l’oreille interne, où elles sont analysées. Ainsi, dans l’audition, deux groupes de phénomènes peuvent être considérés : des phénomènes mécaniques relatifs à la transmission des vibrations acoustiques jusqu’à l’oreille interne, et des phénomènes neurologiques concernant la propagation des influx nerveux vers le cortex. Ils aboutissent au phénomène final : la perception auditive.
Les phénomènes mécaniques
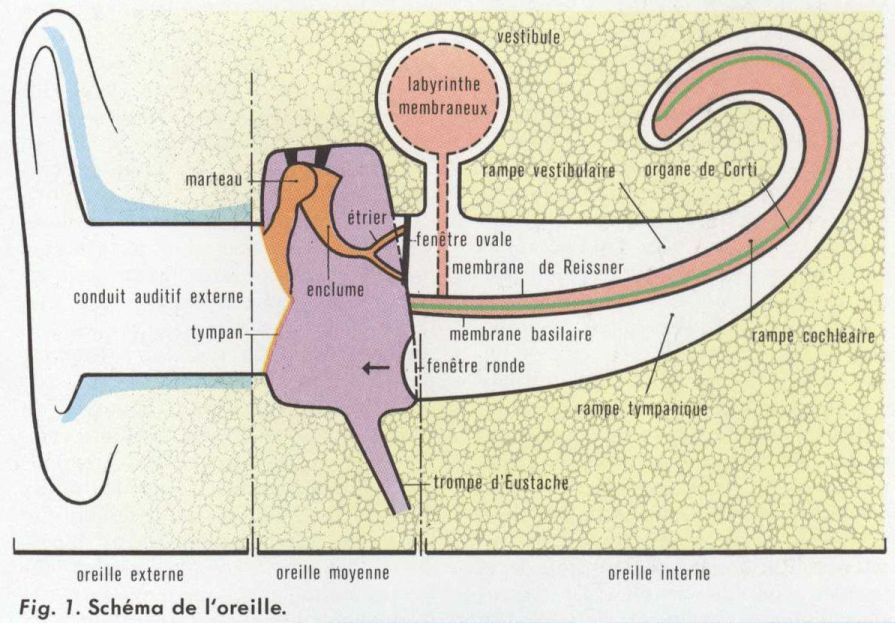
La transmission des vibrations à l’oreille interne se fait par l’intermédiaire de l’oreille externe et de l’oreille moyenne. Le pavillon, par sa forme et sa mobilité chez l’animal, permet de capter les sons et facilite la localisation des sources sonores. Le conduit auditif externe présente une résonance qui amplifie certaines fréquences. L’énergie sonore est transmise favorablement du milieu aérien aux liquides de l’oreille interne grâce à l’augmentation de pression qui résulte de l’action du tympan et de la chaîne des osselets. Des petits muscles, qui s’attachent aux osselets, modifient leur mobilité au cours de leurs contractions. Ils peuvent, dans une certaine mesure, protéger contre les sons trop intenses. Les vibrations de la membrane tympanique et des osselets se transmettent ainsi à la platine de l’étrier. Les mouvements de celle-ci provoquent des variations de pression du liquide périlymphatique (v. oreille) et des autres structures de la cochlée. En raison des caractéristiques mécaniques des membranes cochléaires, les vibrations se répartissent en différents endroits selon les fréquences. Les mouvements qui en résultent agissent sur les cellules sensorielles de l’organe de Corti, où se produisent les processus qui aboutissent à l’excitation des fibres du nerf auditif.
Les phénomènes neurologiques
Les influx nerveux naissent au niveau des fines terminaisons réparties autour des cellules ciliées de l’organe de Corti. À cet endroit se situent des processus de transduction et de codage qui transforment le message acoustique en un ensemble d’influx nerveux, constitués par de petites impulsions électriques qui se transmettent, par le nerf auditif, jusqu’aux centres cérébraux de l’audition.
L’analyse des sons dans l’oreille
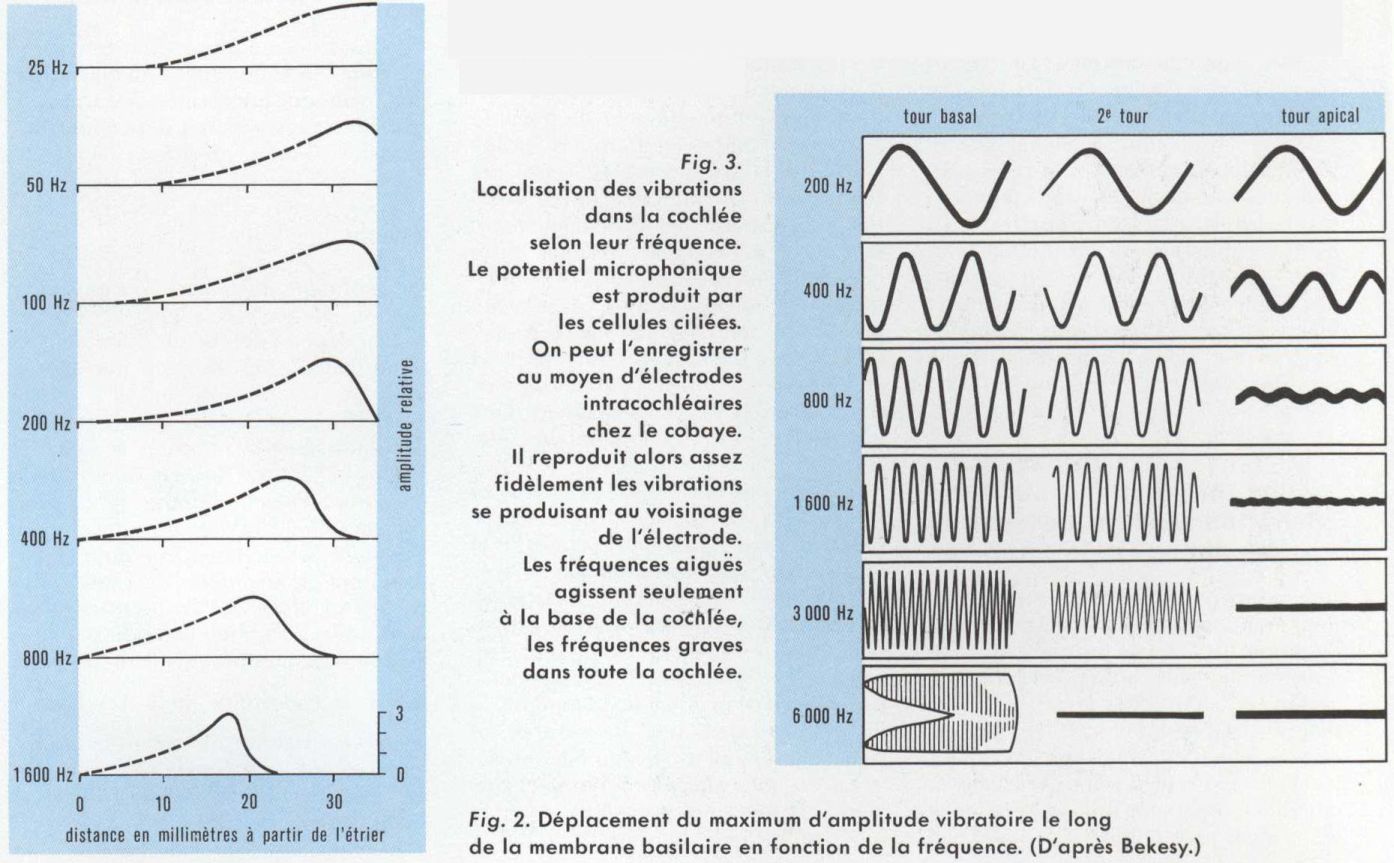
Les vibrations sonores, qui sont transmises par la chaîne des osselets, subissent une analyse à l’intérieur de la cochlée (ou limaçon), grâce à des processus mécaniques qui réalisent une localisation des vibrations le long de la membrane basilaire selon leur fréquence. Schématiquement, la cochlée est analogue à un tube rempli de liquide (v. oreille) périlymphatique. Au milieu, une double cloison délimite le labyrinthe membraneux rempli d’endolymphe. Il prend ici le nom de rampe cochléaire (que l’on appelle aussi cloison cochléaire), et joue un rôle essentiel, car il contient l’organe récepteur auditif, ou organe de Corti. Ce dernier repose sur la membrane basilaire tout le long de la spirale du tube cochléaire. Cette membrane, qui est la plus rigide, détermine en grande partie les propriétés vibratoires de l’ensemble.
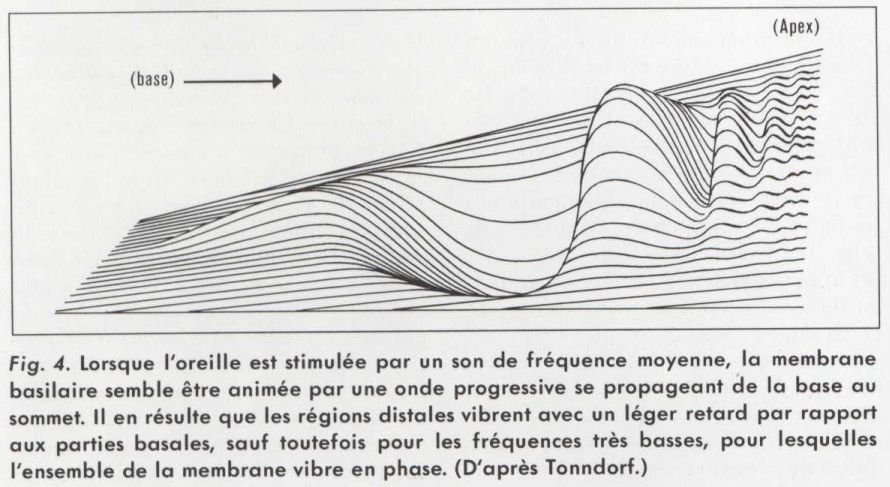
Lorsqu’une vibration sonore pénètre dans la cochlée, grâce au mouvement de l’étrier, le liquide périlymphatique transmet cette vibration aux différentes parties de la rampe cochléaire. Celle-ci présente des propriétés mécaniques variables selon les endroits et des résonances pour les différentes fréquences. Il en résulte que les fréquences basses font vibrer la membrane basilaire sur toute la longueur du tube cochléaire, mais avec une amplitude plus grande vers le sommet. Lorsque la fréquence s’élève progressivement, le maximum d’amplitude se déplace vers la base de la cochlée (du côté de l’étrier). Les parties apicales vibrent alors avec des retards de plus en plus grands, puis cessent de vibrer. Au cours de ces mouvements, l’ensemble de la rampe cochléaire paraît donc animée par une onde vibratoire qui se propagerait de la base vers le sommet (onde progressive). La cochlée se comporte donc comme un filtre peu sélectif.
La localisation des fréquences explique que certaines lésions cochléaires peuvent entraîner une surdité pour une bande étroite de fréquences. Corrélativement, un point donné ne vibre pas pour une seule fréquence, mais pour une bande étroite dite « bande critique ».
Au cours des mouvements de la membrane basilaire, les cellules de l’organe de Corti, ou cellules ciliées, sont déplacées et déformées. Elles sont aussi tiraillées, à leur pôle apical, par la membrane tectoriale. Les déformations ainsi produites provoquent des phénomènes électriques, dont le plus important est le potentiel microphonique, qui reproduit fidèlement la forme de l’onde sonore, comme dans un microphone. Ces phénomènes mécaniques et électriques déclenchent l’excitation des fibres du nerf auditif réparties autour des cellules ciliées.
Cette excitation aboutit à l’émission d’influx nerveux (potentiels d’action), qui se propagent d’abord dans le tronc du nerf auditif, et ensuite, après différents relais, dans les centres cérébraux et dans le cortex. Chaque fibre est excitée par une bande de fréquences qui dépend de l’endroit de la cochlée d’où elle est issue. Ces fibres sont d’ailleurs rangées selon les fréquences qu’elles transmettent.
On remarque que le signal sonore produit dans l’oreille d’abord des phénomènes analogiques (le potentiel microphonique en particulier). Ensuite apparaissent les influx nerveux, qui sont en fait des impulsions électriques comparables à des signaux binaires. Les informations auditives se trouvent donc traduites dans le message nerveux sous une forme codée.
La transmission des informations auditives dans les centres nerveux et les théories de l’audition
Les influx nerveux (potentiels d’action) se propagent dans le nerf auditif et, après des relais (synapses) dans différents noyaux, atteignent le cortex auditif (v. nerveux [système]). Les théories de l’audition s’efforcent d’expliquer comment les informations sont transmises dans les voies nerveuses et comment elles sont interprétées dans les centres pour donner naissance aux diverses sensations auditives. Depuis fort longtemps, des théories ont été proposées pour expliquer la discrimination des divers aspects des stimuli sonores. Un des problèmes qui a suscité le plus de travaux est le mécanisme de la discrimination des fréquences. D’après la théorie de la résonance de Hermann von Helmholtz (1863), chaque point de la membrane basilaire entrait en résonance pour une fréquence particulière et constituait le récepteur spécifique d’une seule fréquence. Les travaux modernes ont confirmé le phénomène de la localisation, mais celle-ci est moins étroite que ne le supposait Helmholtz, en particulier pour les basses fréquences. À l’encontre de la théorie de la localisation, on peut citer les théories dites « téléphoniques » (William Rutherford [1886]), qui envisageaient que la membrane basilaire vibre dans son ensemble de la même manière, comme une membrane de récepteur téléphonique. Dans ce cas, il n’y aurait pas d’analyse, et les fréquences des sons seraient reconnues par le rythme des influx nerveux, qui correspondraient à la fréquence du son. D’autres théories, comme celle d’Ernest G. Wever (1949), tiennent compte à la fois du rythme des influx et de la localisation. Étant donné que les influx nerveux ne peuvent dépasser 1 000 par seconde, seules les basses fréquences pourraient suivre la fréquence du son. Cependant, les fibres peuvent fonctionner de manière alternée, l’une émettant un influx alors que la voisine est au repos, de sorte que l’ensemble des fibres pourrait transmettre des cadences, peut-être jusqu’à 4 000 par seconde (principe de la volée, de Wever). Le phénomène de la localisation ayant été prouvé, on tend à admettre actuellement que la hauteur des sons dépend du rythme des influx pour les basses fréquences, et de la localisation dans la cochlée pour les fréquences élevées.