




respiration (suite)
• Le poumon des Tétrapodes. Son évolution des Amphibiens aux Mammifères (les Oiseaux font exception) est marquée par l’alvéolisation de plus en plus poussée de l’épithélium pulmonaire, qui accroît considérablement la surface respiratoire sans augmenter pour autant le volume de l’organe, et par l’individualisation d’un système conducteur de l’air, d’abord extra-pulmonaire (trachée, bronches), puis intrapulmonaire (bronches, bronchioles, canaux alvéolaires).
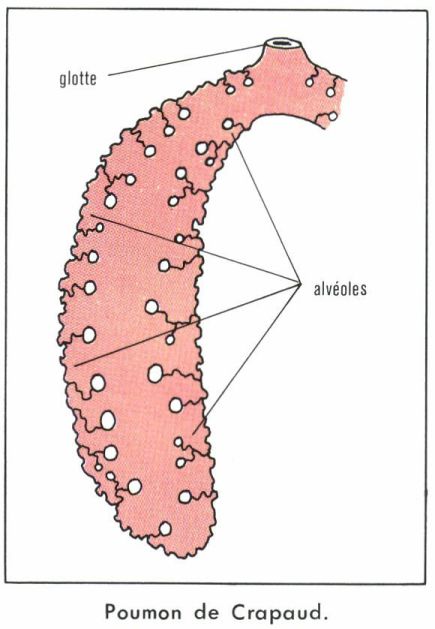
Le poumon des Amphibiens et de nombreux Reptiles est un simple sac à paroi plus ou moins alvéolisée laissant toujours une vaste cavité centrale qui s’ouvre au plancher pharyngien par l’orifice de la glotte, directement (la plupart des Amphibiens) ou par l’intermédiaire d’une courte bronche et d’une trachée commune (Reptiles). En l’absence de cage thoracique, la ventilation pulmonaire des Amphibiens est exclusivement assurée par des mouvements du plancher buccal, qui déterminent des surpressions ou des dépressions assurant l’inspiration et l’expiration. La respiration pulmonaire ne joue qu’un rôle tout à fait secondaire dans les échanges respiratoires des Amphibiens, surtout réalisés au niveau de la peau et de la cavité bucco-pharyngienne (respiration cutanée). D’ailleurs, certains Urodèles, comme l’Hydromanthes du sud-est de la France et du nord de l’Italie, sont dépourvus de poumons. À ces mouvements du plancher buccal, les Reptiles ajoutent des mouvements de muscles thoraciques insérés sur les côtes. Leur contraction dilate la cage thoracique, dont les poumons sont partiellement solidaires, et provoque l’inspiration. Leur relâchement s’accompagne de l’expiration, purement passive.
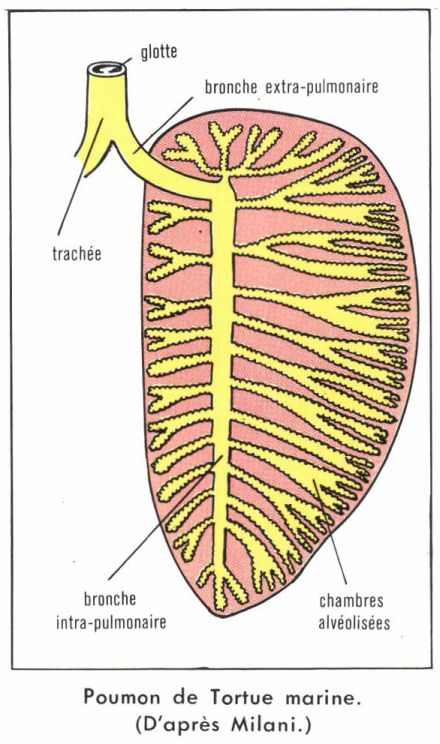
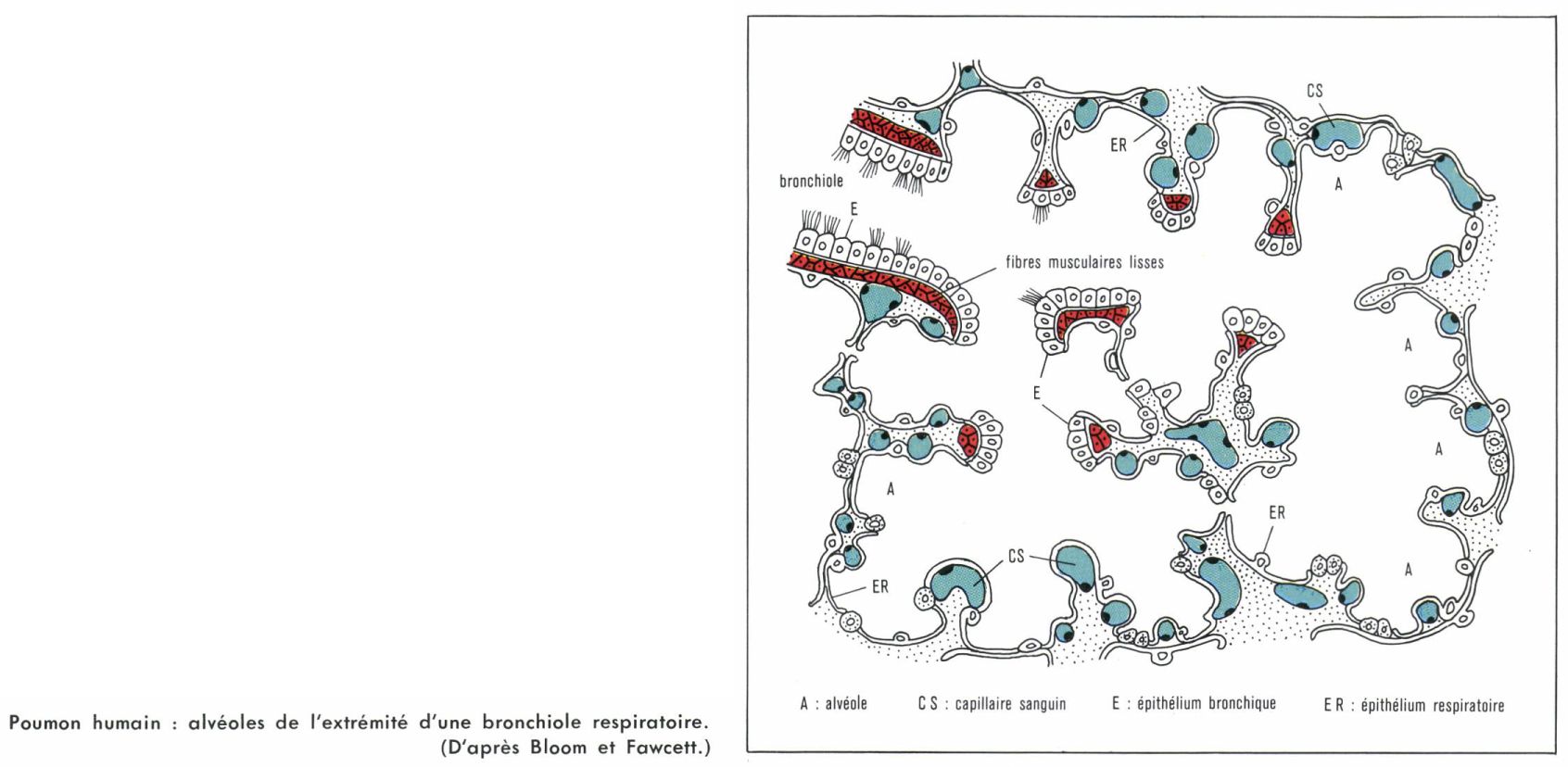
La structure pulmonaire se complique chez les Crocodiles, les Tortues et les Mammifères, par pénétration de la bronche extra-pulmonaire dans la chambre centrale, qui disparaît. Cette bronche intrapulmonaire donne accès à des chambres respiratoires alvéolisées, directement ou par l’intermédiaire de bronches secondaires plus ou moins ramifiées. Le poumon, privé de sa cavité centrale, devient un organe spongieux et homogène. Chez l’Homme, les deux poumons compteraient de 700 à 800 millions d’alvéoles, représentant une surface respiratoire d’environ 90 m2 pour un volume total maximal de 5 litres. La paroi alvéolaire est faite de cellules aplaties, étroitement appliquées contre les capillaires sanguins, de sorte que l’air alvéolaire et le sang des capillaires ne sont séparés que par quelques dixièmes de microns, ce qui permet des échanges gazeux extrêmement rapides.
Les mouvements respiratoires sont assurés, chez les Crocodiles et les Mammifères, par les mêmes muscles thoraciques que chez les Reptiles, mais il s’y ajoute le diaphragme, disposé en dôme entre la cage thoracique et les viscères abdominaux. Sa contraction refoule les viscères vers l’arrière et contribue à agrandir le volume de la cage thoracique et des poumons, qui lui sont solidaires : c’est un muscle inspiratoire.
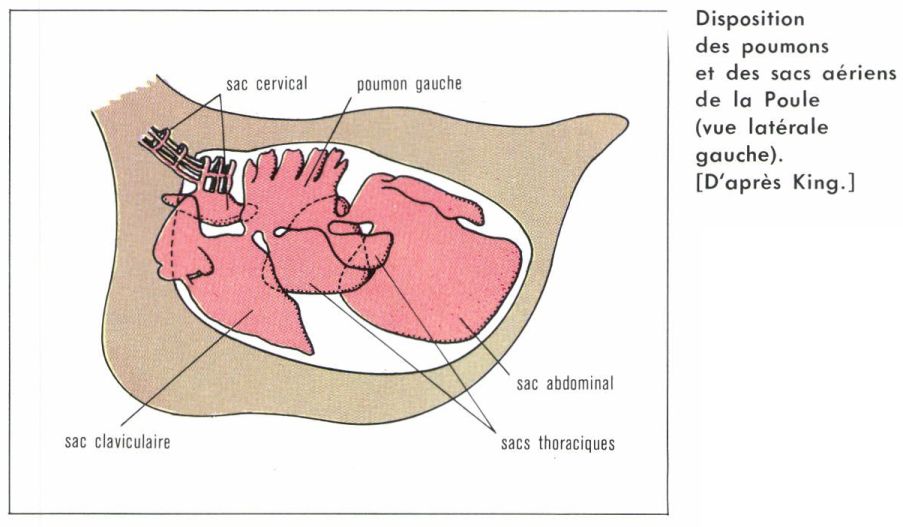
• Le poumon des Oiseaux. Il se singularise par sa structure entièrement tubulaire, sans aucun alvéole, et par le développement de diverticules extra-pulmonaires, les sacs aériens. L’épithélium respiratoire des Oiseaux n’est en effet pas réparti à la surface de culs-de-sac alvéolaires comme chez tous les autres Vertébrés pulmonés, mais au niveau d’un réseau complexe d’anastomoses bronchiques assurant une circulation de l’air pratiquement continue. D’un diamètre de 3 à 15 μ, ces fins canalicules, improprement qualifiés de « capillaires » aériens, sont entrelacés avec un réseau dense de capillaires sanguins. Les sacs aériens sont des diverticules pulmonaires résultant du bourgeonnement de certaines bronches. Ils envahissent la cavité péritonéale, s’insinuent entre les faisceaux musculaires et pénètrent même à l’intérieur de certains os, les os pneumatiques, dont ils occupent la cavité médullaire. En principe au nombre de 6 paires, ils ne sont le siège d’aucun échange gazeux. Ils ne jouent qu’un rôle purement mécanique, en fonctionnant comme des soufflets assurant une circulation continue de l’air à l’intérieur des circuits bronchiques.
La respiration trachéenne
La respiration de la plupart des Arthropodes terrestres (majorité des Arachnides, quasi-totalité des Insectes et des Myriapodes) est assurée par un système de trachées très perfectionné.
Les trachées représentent des invaginations tégumentaires, débouchant à l’extérieur par des orifices pairs à disposition segmentaire, ou stigmates, et se ramifient progressivement dans le corps de l’animal en diminuant de diamètre. Les plus petites trachées (de 3 à 5 μ de diamètre) aboutissent à de volumineuses cellules trachéolaires dans lesquelles elles se terminent par de fines trachéoles intracytoplasmiques de 1 μ de diamètre. C’est à ce niveau que s’effectuent les échanges gazeux. Les cellules trachéolaires et leurs ramifications s’étalent en effet en une sorte de voile sur tous les organes, auxquels elles transmettent directement l’oxygène par diffusion. L’appareil circulatoire, bien que présent, ne joue aucun rôle dans le transport de l’oxygène aux différents tissus, ce qui constitue un cas vraiment exceptionnel.
Chez les Insectes de petite taille, les échanges respiratoires sont passifs et la circulation de l’air dans les trachées résulte d’un simple phénomène de diffusion. Par contre, les Insectes aériens de grande taille et les Insectes à vie active et métabolisme élevé effectuent des mouvements respiratoires, auxquels participent des muscles abdominaux qui compriment les viscères et les trachées et chassent l’air à l’extérieur.

La régulation de la ventilation
Les animaux sédentaires ou peu actifs (Lamellibranches, Vers tubicoles fouisseurs ou parasites) peuvent suspendre temporairement leurs échanges gazeux respiratoires et vivre sans oxygène pendant d’assez longues périodes (30 heures chez les Lombrics, 48 heures chez les Tubifex).