L’ensemble des expériences de résonance magnétique se subdivise en deux grandes classes selon l’origine électronique ou nucléaire des moments magnétiques étudiés. Nous avons évoqué jusqu’ici le cas des atomes, ions ou molécules dont les moments électroniques individuels ont une résultante globale non nulle ; c’est le cas des corps paramagnétiques ; et on parle alors de résonance paramagnétique électronique (R. P. E.).
Mais il existe de nombreux cas où le magnétisme résultant de l’ensemble des électrons est nul ; c’est le cas général pour les molécules. On peut alors observer le magnétisme des noyaux d’atomes, habituellement masqué par le magnétisme électronique parce qu’il est environ mille fois plus faible. On parle alors de résonance magnétique nucléaire (R. M. N.).
Aux moments magnétiques nucléaires mille fois plus faibles correspondent des rapports gyromagnétiques γ et donc des fréquences de résonance ν également mille fois plus faibles. Dans un champ magnétique donné, les fréquences de R. M. N. sont environ mille fois plus faibles que celles de R. P. E.
B. C.
Deux biographies
Sir Joseph Larmor,
physicien irlandais (Magheragall, comté d’Antrim, 1857 - Hollywood, Irlande, 1942). Il a montré que les électrons possédaient une masse et expliqué le phénomène de résonance magnétique grâce au mouvement de rotation de l’atome sous l’action d’un champ magnétique (précession de Larmor).
Rudolf Mössbauer,
physicien allemand (Munich 1929). Il a découvert en 1958 l’absorption par résonance des rayons gamma par certains noyaux d’atomes et le rayonnement qui en résulte (effet Mössbauer). Prix Nobel de physique en 1961.
respiration
Fonction procurant à l’organisme l’énergie dont il a besoin pour vivre, croître et se reproduire, et qui consiste en l’oxydation de molécules organiques à haut potentiel énergétique, qui sont dégradées avec libération d’eau, de gaz carbonique et d’énergie.
La respiration et ses modalités
La respiration cellulaire
Grâce à l’activité photosynthétique des végétaux chlorophylliens, l’énergie solaire (dont tous les êtres vivants sont tributaires) est transformée et accumulée dans les cellules végétales, puis, secondairement et indirectement, dans les cellules animales sous forme de molécules organiques complexes à haut potentiel énergétique (glucides, lipides, protides). La libération de cette énergie nécessaire aux besoins des organismes se produit au cours du phénomène respiratoire, qui oxyde ces molécules et les dégrade en CO2, et H2O notamment. Ces oxydations sont toutefois extrêmement complexes et mettent en jeu des processus originaux qui, s’ils sont bien connus dans leurs principes, nous échappent encore par maint détail.
La phosphorylation oxydative
L’oxydation biologique d’une molécule organique n’a en effet rien à voir avec une simple combustion telle qu’elle peut être réalisée par les chimistes. Ainsi, l’oxydation du glucose ne peut être représentée par la simple équation :
Dans une cellule, cette réaction est décomposée en une cascade d’étapes successives qui constituent ce que les biologistes appellent le métabolisme intermédiaire. Au cours de ces multiples étapes, la molécule organique est progressivement dégradée par oxydation, et de l’énergie est libérée. Cette énergie n’est toutefois pas utilisée immédiatement à la synthèse de nouveaux composés. Elle est provisoirement stockée sous forme de composés phosphatés particuliers, les « phosphates riches en énergie », dont la synthèse nécessite l’addition d’une molécule d’acide phosphorique (ou phosphorylation) et absorbe une forte quantité d’énergie. L’ensemble de ces réactions couplées d’oxydation et de phosphorylation constitue la phosphorylation oxydative.
Alors que la liaison « phosphate » ordinaire nécessite pour se réaliser une quantité d’énergie d’environ 2 calories par molécule-gramme, la liaison « phosphate-haute énergie » (pour laquelle le symbole ~ a été adopté) exige une quantité d’énergie environ 4 fois plus importante. Les phosphates riches en énergie sont des corps thermodynamiquement instables, dont l’hydrolyse rompt la liaison « phosphate riche en énergie » et libère cette énergie. Au premier rang de ces composés se trouve l’adénosine-triphosphate, ou A. T. P., agent universel de stockage et de libération instantanée de l’énergie chez les êtres vivants. Sa synthèse se fait par phosphorylation d’une molécule d’adénosine-diphosphate, ou A. D. P., qui se recharge énergétiquement. Elle a lieu dans les mitochondries et peut s’écrire :
La déshydrogénation et la perte d’électrons
L’oxydation se définit généralement comme une réaction qui consomme de l’oxygène et libère de l’énergie. La réaction inverse est une réduction. Mais l’oxydation (ou la réduction) d’un composé peut se faire d’une autre façon, par libération (ou addition) d’hydrogène. L’oxydation d’un composé peut donc s’écrire : soit A + BO → AO + B + énergie, où A est le composé à oxyder, BO un agent oxydant ou un donneur d’oxygène ; soit AH + B → A + BH + énergie, où AH est le composé à oxyder, B un agent oxydant ou un accepteur d’hydrogène.
Analysés et comparés, ces deux processus d’oxydation (ou de réduction) ne sont que des cas particuliers d’un mécanisme beaucoup plus général d’échange d’énergie, le transport d’électrons d’un atome à un autre. La libération d’énergie est consécutive à une perte d’électrons, qui définit donc une oxydation. En effet, l’addition d’oxygène ou la perte d’hydrogène se traduisent par une perte d’électrons du composé qui s’oxyde et une libération d’énergie. Inversement, l’utilisation d’énergie est consécutive à un gain d’électrons, qui correspond à la définition générale d’une réduction.





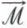 non nulle ; c’est le cas des corps paramagnétiques ; et on parle alors de résonance paramagnétique électronique (R. P. E.).
non nulle ; c’est le cas des corps paramagnétiques ; et on parle alors de résonance paramagnétique électronique (R. P. E.). donné, les fréquences de R. M. N. sont environ mille fois plus faibles que celles de R. P. E.
donné, les fréquences de R. M. N. sont environ mille fois plus faibles que celles de R. P. E.