




population (suite)
Les causes des variations d’abondance
L’intervention de facteurs climatiques abiotiques est évidente dans beaucoup de cas. Les êtres vivants ne peuvent subsister qu’entre certaines limites de température, d’humidité, d’éclairement, etc. Au-delà de ces limites, les populations sont éliminées, et entre ces limites la valeur du taux d’accroissement r varie largement. Autrement dit, les éléments du climat jouent le rôle de facteur limitant, les espèces ayant leur maximum d’abondance lorsqu’elles se trouvent à leur optimum écologique. Ainsi les grands froids réduisent les populations d’Oiseaux ; l’Acarien Panonychus ulmi, qui vit sur les Pommiers dans le nord-ouest de la France, a des populations abondantes après des étés secs et ensoleillés.
La qualité et la quantité de nourriture disponible jouent un rôle important. Les Oiseaux marins sont plus nombreux là où le plancton est plus abondant. Le Grouse d’Écosse, qui se nourrit surtout de Bruyère, recherche les jeunes pousses de cette plante, et leur abondance conditionne le succès de la reproduction de l’Oiseau. Le brûlage périodique de certaines surfaces de lande assure la repousse de jeunes Bruyères et permet au Grouse de reconstituer ses effectifs.
L’intensité de l’action des facteurs que nous venons de citer ne dépend pas de l’abondance des populations : ce sont des facteurs indépendants de la densité. Les facteurs dépendant de la densité ont au contraire une action dont l’intensité est fonction croissante (rarement décroissante) de la densité. Ce sont surtout des facteurs biotiques. Le taux de croissance d’une population décroît avec la densité, soit par diminution du taux de natalité, soit par augmentation du taux de mortalité (l’Homme seul semblant faire exception !). Les raisons en sont fort diverses. Christian et Davis ont montré que des Souris recevant de la nourriture et des abris en quantité suffisante, mais en vivant en milieu confiné, présentent des troubles endocrines croissants qui sont le signe d’une sécrétion accrue de corticostéroïdes ; ces troubles sont dus à des phénomènes de stress résultant de l’agressivité des Souris ; ils conduisent à l’arrêt total des fonctions de reproduction, le retour à la normale ayant lieu quand la densité de population diminue.
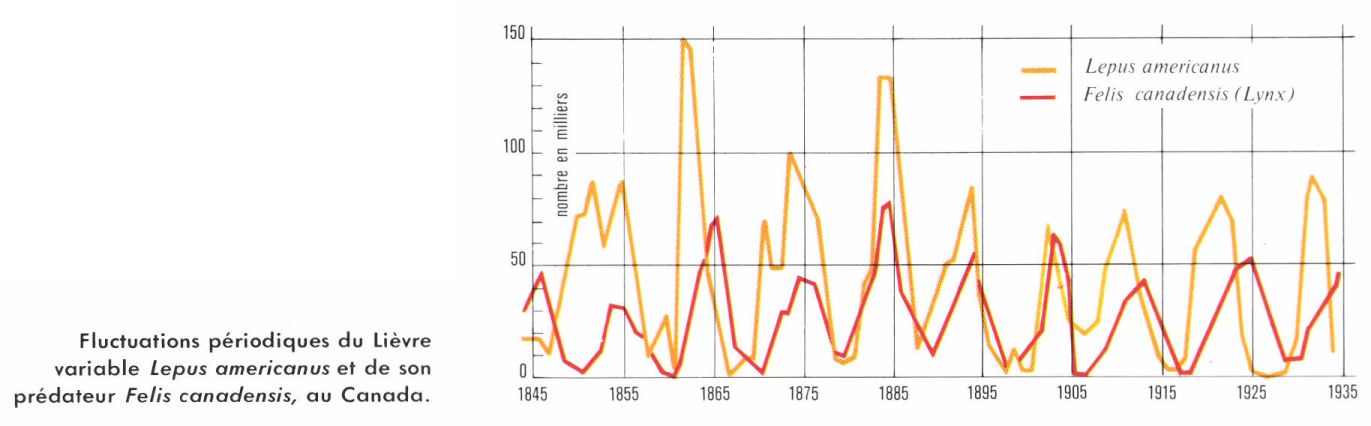
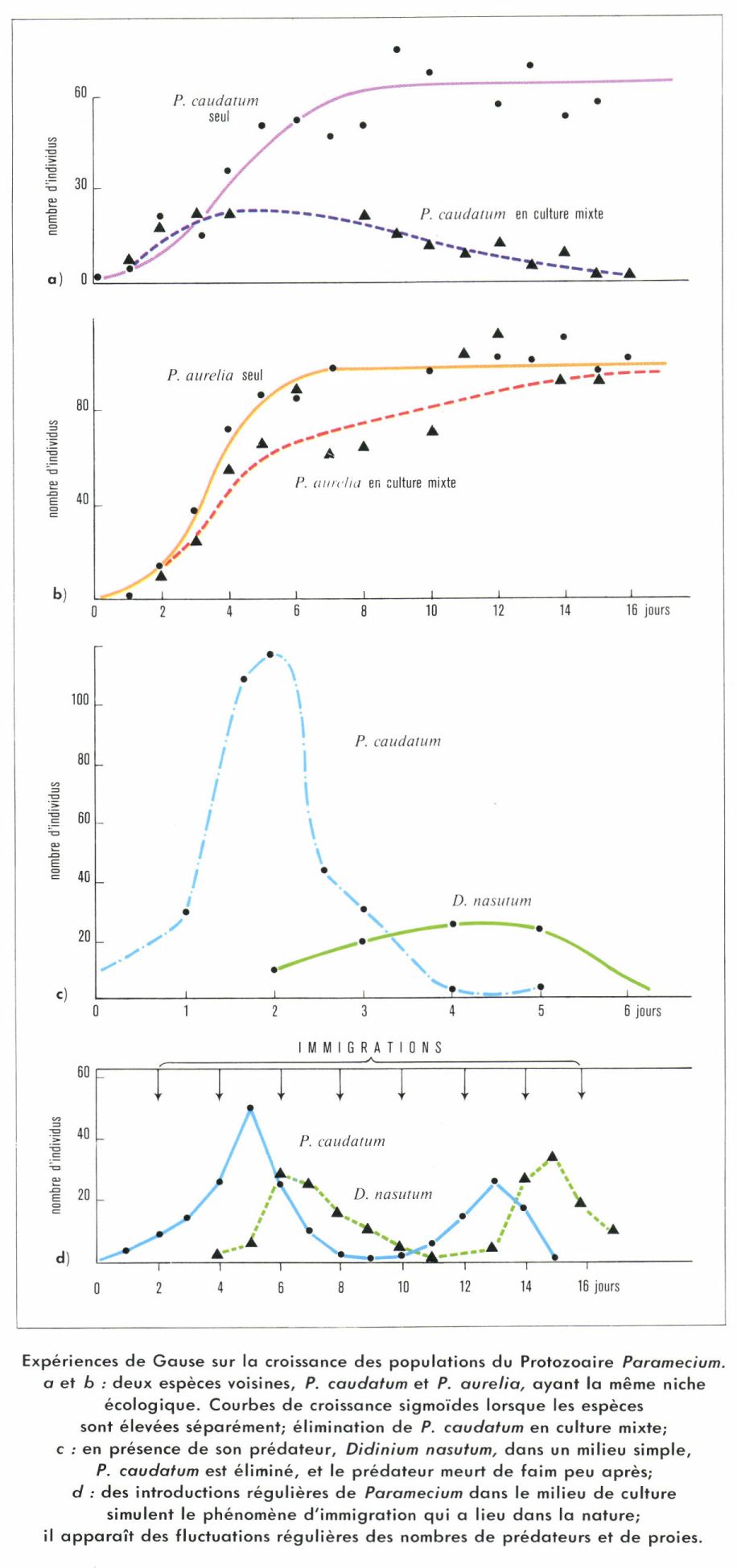
Les parasites et les prédateurs ont une action sur les populations-proies qui est fonction de la densité. Lorsque la proie devient abondante, la fécondité des prédateurs augmente et on peut ainsi expliquer les fluctuations cycliques des Hiboux des neiges et de leur proie, les Lemmings. Les phénomènes de compétition entre individus augmentent aussi avec la densité : dans le cas de la Drosophile, la baisse de la fécondité par femelle est due à ce que les Mouches recherchent les mêmes lieux de ponte et se gênent mutuellement.
La réalité est évidemment complexe, et souvent des facteurs qui dépendent ou non de la densité interviennent simultanément ou successivement pour régler l’abondance des populations. En outre, le rôle des variations génétiques liées à l’intensité de la sélection naturelle et le rôle de l’hétérogénéité du milieu, jusqu’ici à peu près négligés, sont maintenant bien mis en évidence.
L’énumération des théories qui s’affrontent pour expliquer les fluctuations des Lemmings, malgré les très nombreux travaux que ces animaux ont suscités, montrera la difficulté du sujet. Pour certains, la pression sociale consécutive à la surpopulation engendre des phénomènes de stress analogues à ceux qui ont été décrits chez la Souris en captivité. Cela entraînerait une baisse de fécondité et une mortalité accrue en raison de la moindre résistance des animaux aux conditions de milieu défavorables. Une autre théorie admet que la compétition entre individus lors des pullulations provoque une sélection qui est à l’origine de changements qualitatifs favorisant les individus peu féconds. Parfois on fait surtout intervenir des facteurs climatiques. La durée de la période d’enneigement conditionne la durée de la reproduction (les Lemmings ont plusieurs générations par an) et la qualité de la végétation, qui, lorsque la neige tombe tôt, est gelée et conserve sa valeur nutritive en hiver. Enfin, l’intervention des prédateurs est considérée par certains comme fondamentale. Des populations isolées sur de petites îles ne montrent pas de fluctuations alors que sur le continent, à quelques kilomètres, les fluctuations ont lieu en présence de prédateurs. Cette conception concorde donc avec les fluctuations obtenues au laboratoire lors de l’élevage d’un couple prédateur-proie.


La prévision des variations d’abondance et les modèles mathématiques
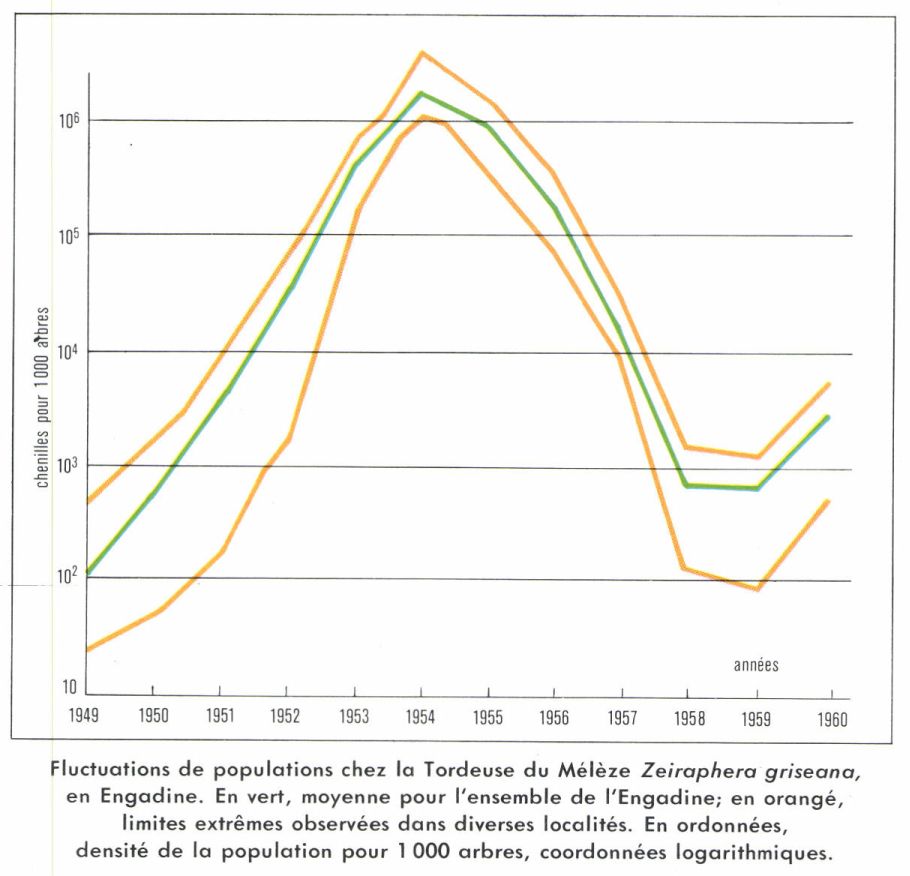
Le but des recherches sur la dynamique des populations est de prévoir l’abondance des effectifs de certaines espèces. Souvent, un seul facteur explique à lui seul une grande partie des variations observées : ce facteur est un facteur clé au sens de Morris. La connaissance des facteurs clés permettra de prévoir en grande partie l’abondance d’espèces nuisibles ou d’espèces utiles. L’établissement de tables de mortalité détaillées fondées sur une étude approfondie de l’espèce est un préliminaire indispensable. Il est alors possible de construire des modèles mathématiques qui ont une bonne valeur prévisionnelle. De tels modèles ont été établis pour divers Lépidoptères ravageurs d’arbres forestiers par Morris pour Hyphantria cunea, au Canada, par Auer pour la Tordeuse du Mélèze en Suisse, etc. Un modèle plus simple pour la Caille de Californie Lophortyx californica, Oiseau gibier très recherché, est dû à Francis. L’analyse faite par la méthode des régressions multiples montre une correspondance étroite entre des paramètres climatiques et l’abondance des populations. Celle-ci a été mesurée à l’aide du rapport du nombre de subadultes (c’est-à-dire d’Oiseaux nés l’été précédent) au nombre d’adultes en automne, lors de la saison de chasse. Les changements annuels d’abondance peuvent être presque entièrement expliqués à l’aide de deux paramètres liés au climat, plus un paramètre lié à la population. L’équation calculée en utilisant les résultats expérimentaux est la suivante :
Q = 0,021 A + 0,929 M – 0,120 P – 0,975,
Q étant la productivité mesurée par le rapport : subadultes/adultes en saison de chasse ; A le pourcentage d’adultes parmi les femelles dans la saison de chasse de l’année précédente ; M la teneur en eau du sol à la fin du mois d’avril et P les précipitations en inches du 1er septembre au 29 avril. L’accord entre les données d’observations et les valeurs calculées est bon et montre la validité du modèle.
Les modèles mathématiques qu’il est possible d’établir à l’aide des ordinateurs sont évidemment empiriques et non explicatifs ; mais ils permettent à la dynamique des populations de déboucher sur des applications pratiques en fournissant la possibilité de prévoir l’abondance des populations étudiées.
R. D.